

Publicado a primeira versão sob Doi: 10.15413/ajsr.2017.0413
https://www.academia.edu/Documents/in/Catastrophic_Geomorphology
Resumo : O paradoxo da estase morfológica na literatura paleontológica, SPM ( stasis paradox morphological ) representa uma grande incógnita que segundo Ernest Mayr, é o maior problema da teoria histórica da Evolução. A SPM é uma anomalia porque esperaríamos encontrar muita diversidade morfológica e taxonômica nas amostras fósseis e nunca repetição de mesmas formas. Se não bastasse este problema , descobrimos outro maior : Fósseis vivos que não mudaram morfologicamente durante centenas de milhões de anos (ficaram em estase) , ao serem submetidos a mudanças ambientais, apresentam mudanças morfológicas hoje e ainda com tendência a não voltar a forma anterior. Ou seja, mesmo se centenas de artigos científicos tentem de várias maneiras justificar a anomalia da PMS e tentar salvar a teoria da evolução histórica, calculamos ser impossível justificarem como fósseis vivos permaneceram sem mudanças em suas várias amostras fósseis e mudarem hoje. Segundo meta-análise de 58 trabalhos de Simpson , existe 71% de repetição de formas nos fósseis, e questionamos este fato com o contraste de podermos assistir em tempo real, mudanças morfológicas geradas facilmente por pressões ambientais. Podemos até fabricar em tempo real mudanças também no nível de sub especiações, gerando inumeráveis variações morfológicas e taxonômicas que são testemunhadas apenas na biodiversidade de hoje, mas não nos trilhões de fósseis e bilhões de amostras . Estimamos haver trilhões de fósseis no mundo, e as bilhões de amostras já coletadas são consideradas como sendo amostras de 570 milhões de anos e até 3,5 bilhão de anos se considerarmos bactérias e fósseis anteriores a explosão cambriana e Ediacara, contando uma historia evolutiva da vida que sai das formas de vida mais simples e acidentalmente vai formando seres mais complexos, porém além de encontrarmos complexidades maiores que as atuais em amostras fósseis antigas, podemos perceber uma tendência maior de simplificação, perda de tamanho, perda de inteligencia, perda de características na historia dos seres vivos e não de ganho, como considerado quando se observa alguma sobrevivência adaptativa. Será que o fato de haver 71% de repetição de formas nos fósseis, indica um catastrofismo global que foi capaz de sepultar as diversas populações de ancestrais tipo básicos defendidos pelo movimento dos biólogos da baraminologia (que defendem os antigos tipos básicos originais da cladística muito citados e "refutados" por Darwin)? Será que o fato da extinção total das famílias e esquemas corporais das primeiras camadas sedimentares contendo fósseis do Ediacara e Cambriano, representam apenas o fato delas estarem mais ao fundo e por receberam maior aporte sedimentar geradas por uma catástrofe global se extinguiram totalmente? Mais estudos são necessários para defender esta mudança de paradigma, porém apresentamos alguns pontos aqui que nos fazem refletir esta possibilidade.
A Extinção das Famílias do Cambriano e Ediacara
A proporção e graduação de extinção se alinha perfeitamente ideia de catástrofe que sedimenta mais, quanto mais fundo for, e portanto extingue mais as famílias do porão dos mares. E Ediacara e cambriano, são típicas formas de seres vivos do fundo dos oceanos. Muitos sedimentos no mar agitadíssimo que foram decantando e preenchendo primeiramente locais mais fundos e por isso extinguiu todos os seres dali. Pois todos os demais repetem em camadas acima, pois numa catástrofe teriam maior possibilidade de não serem submersos vivos por sedimentos. Os que ficam no fundo numa agitação global do mar , tem muito mais chance de serem enterrados vivos que os mais de cima, que tem mais chance de escaparem , fugirem, e serem apenas mortos e arrastados. Uma vez mortos, é mais difícil sua fossilização.

Nível Estratigráfico: Cm = Cambriano; O = ordoviciano; S = Siluriano; D = Devoniano; C = Carbonífero; P = Permiano; Tr = Triássico; J = Jurássico; K = Cretáceo; T = Terciário. Extinção total nível táxon família , aumento de extinções seguido de aumento de sobrevivência em camadas superiores cada vez maior

Imagem indica como era Ediacara
Definições de Espécie, gênero e famílias, e MPTG
Durante décadas, a biologia evolutiva, diante da “plasticidade dos seres vivos” muito citada desde Spencer, 1820-1903 (Lightman & Bernard, 2016), e tão destacada sobretudo desde a imensa revisão bibliográfica feita, quando ainda jovem, por Charles Darwin, tem dificuldades para entender e definir o significado de “espécies”. Atualmente, existem mais de vinte e dois conceitos diferentes de espécies (de Queiroz, 2005). O uso de diferentes conceitos leva a comparações desencontradas na ciência que visa por sistematizar e organizar o conhecimento. Por outro lado, biólogos catastrofistas por décadas têm utilizado para espécies dinâmicas e plásticas, termos e conceitos apropriados mais abrangentes e flexíveis como “tipo” ou “grupo” para o que consideram categorias de organismos relacionados geneticamente, que, em testes de cruzamentos artificiais, gerem emparelhamento cromossômico e pelo menos o início de embrião com características paternas e maternas (Junker & Scherer, 1996). Assim, torna-se prático o agrupamento de clados em torno destes testes de compatibilidade no crossover.
Cada uma das várias categorias de espécies, subespécies e variedades que vemos hoje foram concebidas para se diversificarem de um tipo básico ancestral comum fóssil, portanto consideraremos neste trabalho, a grandeza MPTG (Morphological patterns around the taxon genus), padrões morfológicos em torno de gêneros . Esta grandeza se identifica com fósseis ancestrais, com a falseável hipótese dos tipos básicos (Marsh, 1941), já que “as novas formas tornam-se cada vez mais refinamentos das existentes formas” (Benton et al, 2007). Entre outras observações , podemos dizer que a disparidade morfológica no registro fóssil, quando comparamos táxons mais altos, é alta , e a diversidade de espécies, no registro fóssil, é baixa. Ou seja, um verdadeiro espelho de ancestrais básicos dos quais a maioria de nós (enquanto biodiversidade) descendemos. Analogamente, Muitos troncos e Arbustos com galhos grossos (disparidade), no registro fóssil versus arbustos com infinitas folhagens e raminhos na atualidade(diversidade) . Isso não anula variações morfológicas no registro fóssil , claro que elas existem dentro de um morfoespaço que abriga inclusive mesmos genotipos, e é até esperado que existam evolutivamente , pois as espécies mães possuem cada vez mais , maior pool gênico, uma vez que ainda não estavam desgastadas pelas sucessivas derivas e motores adaptacionais , stressantes e altamente empobrecedores (Mayr), nem pelo acúmulo de genes deleterios que nos lega o motor da entropia genética (Sanford, 2005, 2017; Crabtree, 2010).
Diversos exemplos de disparidade e diversidade são citados na literatura (Benedict et al, 2016). Ou seja, defendemos que MPTG ancestrais básicos em suas características de disparidades morfológicas (e baixa diversidade ) foram sepultados, em geral catastroficamente e com “morte súbita”, para se tornarem fósseis, e este modelo nos conta uma história de um período de um momento catastrófico geológico de: 1) Falta de especiação rápida ou lenta, o que, caso houvesse, enriqueceria a taxonomia fóssil, assim como temos hoje na biodiversidade atual (Benton et al, 2007; Futuyama, 2010) 2) Permanência com alto número de mesmas espécies, o que necessitaria de ambiente estável (repetição fóssil sem pressões ambientais que provocariam manifestações adaptativas e evolutivas), 3) Desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade, tempo longo, e substituiria a parcial sucessão faunística por por “segregação e estratificação espontânea” SEE (Minoletti, 2009, Berthaut, 1986, 1988) que explicam a presença de diversas espécies diferentes segregadas no registro fóssil, bem como não precisam apelar para "ad hoc" para explicarem árvores polistratos e milhares de anacronismos 5) Fósseis de imensos vertebrados completos (o que caracteriza desastre de grande magnitude global e altas taxas de sedimentação, sobretudo de vertebrados completos terrestres), e 6) Mudança drástica no ambiente gerando a radiação adaptativa das espécies nas camadas recentes nas milhões de espécies da biodiversidade atual diversificada por mais que mantenha semelhante numero de padrões corpóreos (Wise, 2013).
Muitas das espécies e suas variações, observáveis hoje, refletem os mesmos padrões morfológicos em torno do táxon gênero (MPTG), apesar de grande parte da literatura designar o táxon superior famílias como padrões morfológicos, o que resulta no fato de lermos na literatura "extinção de famílias "( o que na verdade pode ser apenas gêneros) . Sobre este problema, depois de consultar diversos especialistas e literatura solicitando dados globais sobre quantidade de especies , generos e familias em cada periodo geológico, e receber respostas lacônicas sem solução , comçamos a entender declarações como: “Atualmente, não há uma definição acordada de disparidade, muito menos qualquer consenso sobre como medir” (Wills et al, 1994), então escolhi como melhor síntese a resposta por e-mail, em 2013, encaminhada pelo paleontólogo de Harvard , orientado por Stephen Jay Gould, Dr. Kurt Wise, o qual praticamente repete (porém com maior riqueza) a mesma observação de especialistas consultados, de que as definições e organizações, devido haver inúmeros critérios, estão em falta:
“The best early estimates (by Dave Raup and Jack Sepkoski in the 1980s) were based upon a family-level fossil record data (namely, Sepkoski’s family-level compilation of the marine fossil record). From the number of families that went extinct, the current intra-family species diversity was assumed to be true of the fossil families, and the necessary % species extinction was calculated that would result in that much family extinction (by computer bootstrapping, etc.). Later, after Jack Sepkoski had compiled genus-level data for the marine fossil record (2002, Bulletins of American Paleontology 363), the same sort of calculations were done with the genera that had earlier been done with families. Not only have the species-level data not yet been compiled, but many paleontologists consider that data would be unreliable (because of differences of opinion on how to define paleontological species)”.
Sendo a classificação a nível de famílias algo muito contraditório quando entendemos que família agrupam gêneros que não foram extintos, é comum lermos na literatura dados referentes a uma coisa e outra ao mesmo tempo, ou sobre percentuais ocorridos nas extinção em massa sem o número anterior de onde se retirou tais percentuais. Por mais que isso não tenha gerado tantas correções a nível de descrever extinções (interpretação) ou diminuição de fósseis na estratigrafia (fato), (Sepkoski, 1993), isso inviabiliza ou dificulta outras percepções sobretudo de relação evolutiva no estudo paleontológico. Recentemente gêneros não extintos de bivalves foram classificados como famílias (Gibson, 1996). E devido a isso, tentarmos entender MPTG como uma maneira flexível de lidar com estas imprecisões, o que pode nos ajudar na organização do entendimento geral destas dificuldades classificatórias.
Portanto, MPTG, é o que consideramos como as características peculiares que vão desde os dos tipos básicos matrizes fósseis , até sua descendente diversificação no presente, que ainda hoje estão, exceto famílias e “gêneros” extintos, presentes na maioria das diversas espécies e suas variações encontradas na natureza (Eldredge e Stanley, 1984). Muitas espécies viventes hoje, e suas variações, refletem padrões morfológicos semelhantes. A permanência destes mesmos padrões morfológicos fósseis, por mais que aumente o número de espécies, facilita percebermos os limites da evolução em torno de MPTG , situando e orientando as mudanças possíveis de serem registradas no registro fóssil e testadas seus parentescos na atualidade (Junker & Scherer, 1996). Também o fato desta descontinuidade estar ainda mais marcante no registro fóssil, quanto mais superiores táxons, fortalece esta idéia de descontinuidade sistemática (Carroll, R. L. ,1992).
A tese dos tipos básicos matrizes ou ancestrais (Marsh, 1941), tem sido apoiada pelo fato de o registro fóssil apresentar baixa variabilidade (estase morfológica) e baixa radiação especiativa entre as espécies fósseis (Zimmerman, 1960; Martens, 1997; Albrecht e Wilke, 2008; Alisson, 2013), independente da suas respectivas plasticidades ou maleabilidades fenotípicas ou genotípicas (Ghalambor et al., 2015). Além disso, outros fatores que corroboram a tese dos tipos básicos ancestrais são os mais de 4229 gêneros bem documentados de “fósseis vivos” que são chamados assim por terem sofrido poucas mudanças ao longo do tempo, logo, mantendo-se semelhantes às encontradas no registro fóssil (Romer, 1966; Whitmore, 2013a; Whitmore, 2013b). Provavelmente refletindo idéias de sua época, Darwin chegou a citar essa dedução corrente ao escrever:
"Não há uma verdadeira grandeza nesta forma de considerar a vida, com os seus poderes diversos atribuídos primitivamente pelo Criador a um pequeno número de formas, ou mesmo a uma só? Ora, enquanto que o nosso planeta, obedecendo à lei fixa da gravitação, continua a girar na sua órbita, uma quantidade infinita de belas e admiráveis formas, saídas de um começo tão simples, não têm cessado de se desenvolver e desenvolvem-se ainda!... O venerável e reverendo W. Herbert, mais tarde deão de Manchester, escrevia em 1822, no 4., volume das Horticultural Transactions, e na sua obra as Amaryllidacées (1837, p. 19, 339), que ‘as experiências de horticultura têm estabelecido, sem refutação possível, que as espécies botânicas não são mais que uma classe superior de variedades mais permanentes’. Aplica a mesma opinião aos animais e vê que as espécies únicas de cada gênero foram criadas num estado primitivo muito plástico, e que estes tipos produziram ulteriormente, principalmente pelo cruzamento e também por variação, todas as nossas espécies existentes. Na Nouvelle Flore de l’Amérique Du Nord (1836, p. 6), Rafinesque exprimia-se assim: ‘Todas as espécies podiam ser outrora variedades, e muitas variedades tornaram-se gradualmente espécies, adquirindo caracteres permanentes e particulares’; e um pouco mais adiante (pág. 18) acrescenta: ‘excetuando os tipos primitivos ou ancestrais do gênero’”(Darwin, 1866, capítulo 9, pp.25-26, 577, ênfase nossa).
Quando falamos de estase morfológica no registro fóssil, nos referimos às camadas sedimentares compreendidas entre o ediacara/cambriano até próximo das camadas do período Pleistoceno, ou camadas superficiais, como demonstra diversas publicações baseadas na teoria do equilíbrio pontuado (pontualismo) proposta pelos paleontólogos Niles Eldredge e Stephen Gould (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013). E não nas camadas atuais onde a variação irradia em multiformes variações e ocorre fossilização também.
Os padrões morfológicos ligados ao táxon gênero MPTG, podem ser observados no registro fóssil surgindo repentinamente em todas as camadas sedimentares, com expressivo aparecimento no cambriano e surgimentos abruptos sem nenhum gradualismo em toda coluna geológica, mas uma repetição surpreendente. Essa constatação tem sido divulgada há décadas pelos adeptos do pontualismo que ainda admitem a estase morfológica e a radiação florescendo em especiações somente nas camadas geológicas mais atuais (Levinton e Chris, 1980; Woodruff, 1980; Williamson, 1981; Eldredge, 1986; Van Bocxlaer e Hunt, 2013). Estudo evolutivo recente corrobora tal afirmação ao dizer que :
"A visão dominante da evolução baseada no registro fóssil é que as espécies estabelecidas permanecem mais ou menos inalteradas durante sua existência. Por outro lado, a evolução substancial é rotineiramente relatada em populações contemporâneas, e a maioria dos traços quantitativos mostra um alto potencial de evolução. Essas observações contrastantes em escalas de tempo longo e curto são muitas vezes referidas como o paradoxo da estase, que se baseia na suposição fundamental de que os períodos de estase morfológica no registro fóssil representam uma mudança evolutiva mínima." (Voje, 2016).
Assim, o presente artigo tem como objeto de estudo esses fatos observados e bem documentados: surgimento repentino fóssil, estase morfológica, repetição de mesmas espécies fósseis em contraste com a explosão de radiação adaptativa revelando todo potencial de plasticidade e maleabilidade dos fenótipos e genótipos dos seres vivos que deveriam estar presente no registro fóssil caso a história evolutiva tivesse ocorrido ali, pois “a diversidade morfológica diminui juntamente com a diversidade taxonômica. Este padrão sugere heterogeneidades como a extinção elevada e / ou a origem reduzida em certas regiões do morfoespaço” (Foote, 1993).
As especiações e variações que, como vimos, carregam os MPTG podem ser observadas ocorrendo em tempo real e histórico-arqueológico, mas estão ausentes na mesma proporção, no registro fóssil. Isto se torna relevante na medida em que percebemos o sucesso reprodutivo das variações e sua permanência ao longo do tempo, pois a variação em torno de gênero não exige grandes bio-transformações, mas deve ocorrer de qualquer forma, em torno do gênero (MPTG) nas amostras fósseis. Hoje, surpreendentemente, segundo o paleontólogo especialista em estatísticas fósseis Dr. Kurt Wise, tecnicamente, o número de planos corporais no registro fóssil não excede o número de planos corporais hoje (Wise KP. 1989), e segundo esta afirmação, temos quase o mesmo número de padrões corporais entre os 250-300.000 fósseis catalogados, comparados aos 2 milhões de espécies vivas hoje já catalogadas, com estimativas de 8,7 milhões e estimativas que vão até 100 milhões de espécies hoje) e as ínfimas 300 mil espécies fósseis do cambriano/edicara ao pleistosceno. (Woodmorappe, 2000; Sadava et al., 2009; Mora et al., 2011; Catalogue of Life, 2016). Quando comparados os padrões morfológicos, esperar-se-ia que tivéssemos um número bem maior de PMTG no registro fóssil se este espelhasse uma amostra da biodiversidade ocorrida em 544 milhões de anos correspondentes ao Fanerozóico. Portanto, o fato do número de espécies atuais, na casa dos milhões, não conseguir transpor o número de padrões corporais presente no registro fóssil, sugere evolução delimitada a padrões morfológicos ou tipos básicos ancestrais (baraminologia) ou MPTG.
Especiação em Tempo Real
A surpresa de muitos diante da especiação rápida e mudanças significativas morfológicas em apenas uma geração, refletia a falta de previsão da teoria do que se esperaria em termos de tempo. Entendemos a especiação em tempo real como um fenômeno em que duas ou mais populações de uma mesma espécie, se modificam em novos arranjos de informação genética pré-existente, tendo como causa a separação por barreiras geográficas e mortandades, porém em um tempo hábil para que seja possível a observação de todo o processo do início ao fim. (Furness et al 2015). Experiências empíricas promovendo especiação em tempo real , tem sido testadas com sucesso e colocando em xeque cada vez mais o PMS, (Ghalambor et al, 2015), o que demonstra o que “nenhum organismo ignora seu ambiente” (Ezard et e al, 2016).
Podemos ainda classificar o processo de especiação de duas formas: em tempo real ou em tempo histórico-arqueológico. A especiação em tempo real é aquela em que ocorre bio-modificações limitadas ao tipo básico, equivalentes ao surgimento rápido de uma nova “espécie”, observada ̶ por experimentação ou observação não-controlada ̶ em até 50 anos. A especiação em tempo histórico, por sua vez, diz respeito às bio-modificações equivalentes a uma nova “espécie” acima de 50 anos que podem ou não ser observadas devido o tempo de vida do observador ou projeto de pesquisa. Nesses casos, há 50% de chance de observação e os outros 50% restantes dependem do uso da dedução, por exemplo, a partir de análise cromossômica e cálculos de taxas de radiação (Trivedi, 2000).
O papel da deriva genética no processo de especiação
Existem diversos mecanismos envolvidos no processo de especiação (epigenética, deriva genética, seleção natural, influências ambientais e geomagnéticas no crossover (Gorelick, 2005), etc). Devido à especiação estar intimamente relacionada à deriva genética e a conseqüente perda de pool gênico de uma população anterior, estudos do modelo de muitos ancestrais básicos (SRABUC) que apostam na especiação em tempo real, se tornam fundamentais para explicar as evidências atuais de limites à adaptação nos diferentes organismos vivos (Bell, 2013).
A deriva genética é um mecanismo que modifica aleatoriamente e de forma repentina – devido a catástrofes ou a isolamentos diversos, por exemplo, − as frequências alélicas de uma população (Ridley, 2006; Freeman e Herron, 2009). Por outro lado, acredita-se que a seleção natural é direcionada, isto é, elimina muitas mutações deletérias, mutações necessárias se perdem juntas, ignora as mutações neutras e deletérias (principalmente as que se manifestam após período reprodutivo) seleciona os traços sobreviventes e/ou vantajosos pré-existentes ou que comunicaram em face de resposta epigenética ao meio (Sanford, 2014). Porém, como afirmou a bióloga americana Lynn Margulis, “a seleção natural elimina, e talvez mantenha, mas ela não cria” (Teresi, 2011, p. 68). Ademais, devido o fato da deriva genética não fazer distinção entre genes bons e ruins, e a seleção natural falhar também em seu filtro, isso resulta em perda de variação genética (Lacy, 1987), deixando as espécies geneticamente mais pobres e “cada vez mais próximas da extinção”. Em outras palavras, a deriva decepa e empobrece a população de seu “pool gênico” e a seleção natural ajeita mais ou menos o que restou.
Nesta frase farei uma modificação, pra que fique mais técnico.Sabe-se que o efeito da deriva é inversamente proporcional ao tamanho da população, podendo aparecer em diferentes momentos da história das espécies e da própria humanidade. Nesse sentido, é possível que a deriva tenha tido um papel fundamental e sobrepujante ao da seleção natural no que diz respeito ao processo de especiação rápida após uma grande catástrofe.
O processo de especiação após Catástrofes
Grandes catástrofes mundiais são as formadoras da maioria das camadas sedimentares presentes no globo, que estão associadas a imensas inundações e destruições em todo globo (Souza Jr, 2008). As conseqüências da mortalidade em massa gerada, geraram stress endogâmico nas espécies sobreviventes que quanto mais se afastassem mais isolamentos geográficos geográficos e derivas gerariam explicando assim a radiação recente fabricando toda a biodiversidade super variabilizada de hoje , que está contrastada com a pobreza morfológica e palpérrima ou nula manifestação sub especiativa do registro fóssil.A mudança do planeta alem de outros fatores, geraram condições muito favoráveis para uma rápida especiação (Wilmer et al., 2011). O modelo criacionista defende o sepultamento da biodiversidade de todo planeta por estratificação espontânea nas transgressões e regressões marinhas (Berthault, 1986; Berthault, 1988; Brand e Tang, 1991;Snelling, 1997; Berthault, 1998; Chadwick e Spencer, 2006; Berthault, 2013), seguido de especiações rápidas pelos motores evolutivos adaptacionais que diversificariam os padrões de ancestrais prontos sepultados em um catastrofismo inédito .
Nesse contexto, a especiação em tempo real legitima o modelo de um “salto evolutivo” recente, com dois picos bio-diferenciadores relacionados aos episódios do inicio da entropia genética humana (Fu et al., 2013) depois das catástrofes associadas que empobreceriam o pool gênico dos seres vivos e dariam aos sobreviventes, em virtude da migração e do isolamento geográfico, o legado de efeito fundador – situação frequente na especiação peripátrica− em um planeta com um ecossistema totalmente reconfigurado onde o motor epigenético atuaria para corresponder as necessidades do novo ambiente (Ridley, 2006; Eakin, 2014; Weyrich et al., 2016).
A especiação peripátrica é um mecanismo pelo qual podemos explicar o enorme aumento da diversificação pós-catástrofes. É um tipo de especiação pelo qual as espécies novas são formadas em populações periféricas isoladas (Ridley, 2006). Na especiação peripátrica, populações drasticamente reduzidas fazem com que a especiação completa seja o resultado mais provável do isolamento geográfico, porque a deriva genética age mais rapidamente em populações pequenas. Deriva genética somada a fortes pressões seletivas, causariam uma rápida mudança genética na pequena população descendente (Wilmer et al., 2011).
Observações atuais podem servir de exemplo para compreendermos melhor os casos de especiação peripátrica. Estudo recente afirma que “uma análise de mais de 2.000 espécies de pássaros fornece uma visão sobre como evoluíram as diversas formas de bico dos animais e aponta para um único evento raro como um gatilho para a rápida divergência inicial das linhagens aviárias.” (Bhullar, 2017).
Toda essa sucessão de fatos pode ser vislumbrada por meio de um cenário onde a maioria dos organismos destruídos por essas grandes catástrofes associadas e consequentes, deixaria pequenas populações de sobreviventes (Wilmer et al., 2011). Segundo matéria publicada pela Folha de São Paulo a respeito deste achado, “o que pareceu fascinar mais outros biólogos, porém, é a grande velocidade com que o fenômeno do deslocamento de caráter ocorreu. ‘Eu acreditava que fosse demorar muito mais’, comentou na ‘Science’ o biólogo David Pfening, da Universidade da Carolina do Norte. A redução média de 5% no tamanho de bico, considerada drástica pelos biólogos, ocorreu no intervalo de cerca de um ano, praticamente de uma geração para a outra.” (Folha de S. Paulo, 2006).
A surpresa dos biólogos evolutivos com a descoberta desse novo fenômeno (Grant, 2017) se dá devido à falta de entendimento de outros motores coadjuvantes, que atuam nessas bio-modificações limitadas ao tipo básico, tais como epigenética e aspectos energéticos e de temperatura atuando no crossover (Fondon e Garner, 2004; Eakin, 2014). Para a Dra. Jean K. Lightner,45 parece haver três fontes para as variações associadas a radiação adaptativa: 1) hibridização, 2) mutação e 3) triagem ambiental de alelos ancestrais (por mecanismos de seleção natural e deriva meiótica) (Lightner, 2016).
Estas observações, se não encarecem a dívida dos elos perdidos na paleontologia pontualista, aumenta muito a dívida de variabilização taxonômica fóssil para o modelo histórico geológico que assume pelo menos cinco grandes catástrofes separadas por milhões de anos na história da terra.O curioso é que as pesquisas ligadas a especiação em tempo real tanto confirmam as observações de Darwin no âmbito da biologia funcional (equilíbrio pontuado) quanto destrói por completo os postulados evolutivos em termos de períodos geológicos (gradualismo filético), e é perfeitamente comunicável a modelos catastrofistas que aposta numa especiação rápida impulsionada pelo efeito das bio-modificações limitadas ao tipo básico.
Baraminologia e o estudo dos “tipos básicos”
O aparecimento repentino e pronto de formas de vida e de sistemas irredutivelmente complexos (Lonning, 2005), além do número tecnicamente igual (Wise, 1989) de padrões morfológicos quando comparamos taxonomia fóssil de 250-300 mil espécies com estimativas de 9 a 100 milhões atuais, sugere a idéia de um fenômeno de tipos básicos ancestrais surgidos prontos e a permanência numérica de padrões corporais de uma evolução limitada em torno deles (MPTG). Há uma comunicação muito fluente entre o darwinismo biológico e o modelo SRABVSURCbiológico, onde muitos se valem de descobertas evolutivas, como especiação em tempo real, para se aproximar mais da historicidade de curto período de tempo, e utilizando a radiação recente como justificativa para toda a biodiversidade. A teoria da evolução sintética, mais especificamente atuando no âmbito biológico, que ensina a diferenciação das espécies, neste contexto, tem baixa discórdia entre as classes mais avançadas de SRABUC, sobretudo nos cientistas do movimento de “Baraminologia” (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). Suas discordâncias se concentram mais no âmbito da história geo-paleontológica compreendida entre o cambriano/edicara e pleistoceno e/ou camadas superficiais.
Desde 1941, o modelo SRABVSURC é contrário a ideia de que especiação é sinônimo de “evolução” por meio de estudos relacionados às especiações no campo da Baraminologia (Marsh, 1941; Wise, 1992; Robinson e Cavanaugh, 1998; Frair, 2000; Jerlström, 2000; Wood e Cavanaugh, 2001; Cavanaugh e Wood, 2002; Wood, 2010; Aaron, 2014). O biólogo norte-americano Dr. Frank L. Marsh, um dos fundadores da Creation Research Society foi quem cunhou a palavra "baramin". (Marsh, 1941; Frair, 2000) Ela foi derivada da combinação de duas palavras hebraicas − bara (criado) e min (tipo) – referindo-se a tipos básicos criados (espécies, em versões bíblicas em português) (Frair, 2000).
Em 1990, o paleontólogo catastrofista Kurt Wise observou a necessidade de uma biossistemática SRABVSURC− um método de estudo, nomeação, e classificação de baramin (Wise, 1990; Frair, 2000) ou ancestrais de MPTG como defendemos aqui. O campo científico foi oficialmente denominado como “baraminologia”, que de forma simplista significa o estudo dos baramins ou dos tipos básicos ancestrais. Segundo os pesquisadores Reinhard Junker e Siegfried Scherer,“tipos básicos é uma unidade de classificação, um taxon, resultado do trabalho da descontinuidade sistemática como é observado na natureza” (1996, p.34; Wood et al., 2003). Dito de forma simples, tipos básicos criados variabilizaram ao longo do tempo até chegarem ao que conhecemos hoje como subespécies.
Existem algumas regras falseáveis para se considerar um grupo de espécies como pertencentes a um tipo básico ancestral comum. O Dr. Junker e o Dr. Siegrifield Scherer destacam na 6ª edição alemã do livro Evolução, um livro texto crítico:
“todos os indivíduos que estão unidos direta ou indiretamente por cruzamentos são considerados pertencentes a um tipo básico (nível genético). E todas as espécies biológicas que se se assemelham claramente umas as outras pertencem a um gênero (nível morfológico). E todas as espécies biológicas que em princípio podem cruzar entre si pertencem a um tipo básico (nível morfo-genético)”... “dois indivíduos pertencem ao mesmo tipo básico quando a embriogênese de um híbrido vai além da fase maternal do desenvolvimento e contem uma expressão coordenada e genes morfogenéticos paternos e maternos” (Junker e Scherer, 1996, p.34).
Além disso, os baraminologistas usam uma série de critérios metodológicos de adesão para determinar os limites dos grupos baramins (Robinson, 1997; Wood, 2001; Wood, 2002; Wood, 2005; Cavanaugh e Sternberg, 2004). De modo geral, os métodos mostram espacialmente graus de similaridade e de dissimilaridade entre grupos, e podem revelar informações taxonômicas úteis, distinguindo cada vez mais os fatores que dão probabilidade ou não de parentesco, aumentando assim sua contribuição em biologia aplicada a técnicas de melhoramento genético e estudo do comportamento evolutivo das populações.
A baraminologia, também conhecida como sistemática de descontinuidade, está rapidamente se tornando uma das áreas mais ativas de pesquisa SRABVSURC (Scherer, 1993), e algumas de suas metodologias têm sido aplicadas e testadas até mesmo por pesquisadores ligados ao modelo gradualista geológico de ancestralidade totalmente comum com destaques em periódicos com peer-review (Senter, 2010; Wood, 2011). Como já vimos, sua principal finalidade é determinar quais organismos compartilham um ancestral comum (Frair, 2000). A ideia básica defendida neste campo de pesquisa é a de que há limites nas possibilidades de cruzamento que não podem ser atravessados. Nesse contexto, baraminologistas objetivam encontrar as "descontinuidades" na história de vida, ou os limites da ancestralidade comum (Remine, 1993).
Esse campo de pesquisa ganha ainda mais estímulo frente às evidências atuais mostrando que “especialistas” têm erroneamente classificado algumas espécies dentro de um dado gênero por conta do “desejo” em descobrir o ancestral comum universal (Lopes, 2015). Paleontólogos afirmam que um terço das “espécies” reconhecidas como sendo de dinossauros talvez nem mesmo tenha existido (Horner e Goodwin, 2009). Para eles, essas “espécies” podem não ser espécies separadas, mas estágios juvenis ou subadultos, em desenvolvimento, identificados erroneamente como exemplares de outras espécies. Em artigo publicado na revista Science, por exemplo, Schwartz e Tattersall afirmam que esse milagre da multiplicação da nomenclatura das espécies foi longe demais (Schwartz e Tattersall, 2015).
Vale lembrar que, embora a Baraminologia tenha alcançado resultados promissores, suas conclusões não são definitivas (Wilson, 2010). Por ser um campo recente, mais pesquisas são necessárias e seus métodos e técnicas recém-elaborados devem ser mais bem examinados a fim de legitimar ou não a sua função e utilidade na caixa de ferramentas da ciência.
Entropia genética na especiação
E o que dizer dos custos causados quando realmente ocorre a especiação? Outro grande dificultador para o modelo gradualista é que ele é neutro em relação à melhora ou piora do processo de especiação, embora assuma que a deriva genética cause perdas de variação genética em populações pequenas. O modelo de muitos ancestrais básicos sepultados num catastrofismo recente (SRABUC), por sua vez, munido da proposta da entropia genética, defende que a especiação resulta na perda de informação genética e na consequente degeneração do genoma devido às adaptações como recursos evasivos, perigosos, desgastantes para as populações (Ariza, 2007; Sanford, 2014; Crabtree, 2013a; Crabtree, 2013b). Outras evidências corroboram o modelo SRABVSURCno sentido de sugerir que essa perda de informação genética devido a mutações deletérias em humanos tenha se dado recentemente, entre 5.000 a 10.000 anos atrás (Fu, 2013).
A entropia na informação genética é cada vez mais evidente em tudo que se observa: na deriva genética, na seleção/eliminação, nas mutações, nas complexidades imunológicas desgastantes, entre outros mecanismos. Em humanos, por exemplo, as estimativas atuais é a de que ocorram entre 100-200 novas mutações por indivíduo a cada geração (Nachman e Crowell, 2000; Dolgin, 2009; Lynch, 2010). Destas, os dados variam entre 1-15% de mutações deletérias que causariam a perda direta de informação genética em humanos a cada geração (Nachman e Crowell, 2000; Lynch, 2010; Eyre-Walker e Keightley, 1999;Shabalina et AL., 2001; Keightley, 2012).
Em relação ao fitness, um estudo publicado em 1997 estimou entre 1-2% a taxa de perda da aptidão humana, ou seja, um custo genético alto que faz com que a humanidade se degenere a cada geração devido o esgotamento dos recursos adaptativos utilizados para manter sua variedade genética (Crow, 1997). Em 2010, por sua vez, outro estudo estimou que a aptidão humana está em declínio em 3-5% por geração (Lynch, 2010). Também, outro estudo simulando numericamente o acúmulo de mutações deletérias, demonstra além de a maior parte das mutações estarem a esquerda, (-0.001), ou seja, tendência de serem deletérias, “mutações ruins”), elas também estão acumulando, em uma zona chamada de “Zona de Não-Seleção”, por serem “quase-neutras” essas mutações vem se acumulando ao longo das gerações causando dezenas de “erros ortográficos” no genoma populacional. (Sanford et al 2008)
Figura 2 - Distribuição simulada do acúmulo e frequência de mutações deletérias ao longo de gerações e seus efeitos na aptidão populacional.

Gráfico em vermelho mostra distribuição natural de mutações onde não ocorre seleção. As colunas em verde, mostram as mutações atuais que estão se acumulando, ou seja, dominantes. Outras colunas são mutações menores, recessivas que vem se acumulando à medida que chegam a zona quase neutra, próxima à 0.0.
Sucessivas Derivas Empobrecedoras
O zoólogo holandês Duyvene de Wit descreveu perfeitamente esse processo de empobrecimento genético ao afirmar que:
“quando uma população marginal abre caminho para um novo habitat, ela não pode levar consigo todos os genes da população materna, mas sim apenas parte deles. Cada nova raça ou espécie que se origina de outra possui, portanto, um pool gênico mais pobre. Conseqüentemente, a perda de substância do ‘pool’ gênico é o preço que cada raça ou espécie tem de pagar pelo privilégio de existir. Se o processo de especiação ocorrer repetidas vezes consecutivamente, surgirão por fim, espécies cujos ‘pools’ gênicos estão tão completamente esgotados que bastam alterações insignificantes nas condições ambientais para que elas venham a se extinguir. Esforços para se adaptar a modificações ambientais como resultado de possibilidades insuficientes de recombinação levam, por fim, a um estado genético mínimo. Se este limite mínimo for ultrapassado, não haverá mais possibilidade de sobrevivência. Por essa razão, o destino trágico e irrevogável das espécies ou raças especializadas, extremamente adaptadas, é a morte genética” (Kahle, 1999, p.87; Junker e Scherer, 1996).
Portanto, a proposta SRABVSURC é razoável e se torna cada vez mais corroborada pelos dados científicos ao afirmar que os seres vivos do passado eram mais flexíveis evolutivamente , tinham maior fitness adaptativo, eram saudáveis e sem acúmulo de carga negativa de mutações deletérias e não eram aptos em relação aos problemas de hoje, eram sem os defeitos oriundos das adaptações forçadas e das pressões seletivas distintas a fim de que viessem a subsistir. Vale lembrar que sub-especiações, quanto mais verificáveis em tempo real, mais provam a bio-modificação em curto período de tempo, tal como prevê o presente modelo neocatastrofista de impactos de asteroides.
Registro fóssil e a ausência de diversificação de espécies
Especiação proporcional ao comportamento biodiferenciador observado está ausente pelo fato de existir apenas cerca de 250-300.000 tipos fósseis se repetindo dentre as estimativas de trilhões ou inumeráveis amostras, em todos os estratos geológicos até o pleistosceno (Woodmorappe, 2000; Sadava et al., 2009; Loceye Lennon, 2016). Este fato indica a ausência de sequências durante o processo de especiação quando contrastada com as estimativas de 8,7 milhões de espécies vivas atualmente (Mora et al., 2011). Este salto indica uma catástrofe no meio , assim como temos a obviedade de inicio de camadas sedimentares trazendo rica informação fóssil no ediacara/cambriano e todo o diferencial das camadas do fanerozoico com as pré-cambrianas, tudo nos fala de uma gigantesca catástrofe que iniciou sua deposição ali .
Em outras palavras, a ausência de “diversidade” no registro fóssil revela que as diferenciações ocorrem mais no presente (milhões de espécies) do que em toda coluna geológica (com apenas 3 centenas de milhares). Este episódio indica um tempo em que os seres vivos não precisavam se sub-especiar (bio-modificar) e se adaptar com frequência pra sobreviver, porque viviam em um ambiente mais propício para a vida, estável, assim como a PMS requer.
Evidências recentes apontam que o início da diversificação de alguns gêneros de plantas consideradas “fósseis vivos”, por exemplo, ocorreu ao mesmo tempo ao redor do mundo e em um período muito mais recente do que se supunha, revelando conflitos na proposta gradualista das espécies (Nagalingum et al., 2011). Ademais, foi relatado que essa rápida diversificação teve como causa uma grande mudança climática. Outro exemplo semelhante vem de Ernest Mayr :
“In evolutionary biology we have species like horseshoe crabs. The horseshoe crab goes back in the fossil record over two hundred million years without any major changes. So obviously they have a very invariant genome type, right? Wrong, they don't. Study the genotype of a series of horseshoe crabs and you'll find there's a great deal of genetic variation. How come, in spite of all this genetic variation, they haven't changed at all in over two hundred million years while other members of their ecosystem in which they were living two hundred million years ago are either extinct or have developed into something totally different? Why did the horseshoe crabs not change? That's the kind of question that completely stumps us at the present time.”
Aliás, são vários os motores adaptacionais e modificacionais integrados reconhecidos atualmente. O fato de eles existirem nos seres vivos, nos permite sugerir que as bio-modificações nunca deixaram de existir, nunca e em hipótese alguma não podemos dizer que evolução e flexibilidade nos seres vivos é fato somente hoje.
Outro padrão do registro fóssil parece sugerir mais o movimento de segregação e estratificação espontânea que (Dilly et al, 2015: Berthaut, 1986, 1988, 2002, 2004, 2010, 2011, 2012, 2013, 2014; Lalomov, 2007, 2013; Julien, P.Y, 1993), reproduzem em laboratório . Um deles é o desaparecimento coordenados: “Desaparecimentos coordenados . Um grande número de espécies fósseis podem desaparecer do registro geológico em um nível estratigráfico específico (ver Figura 3). O desaparecimento nunca está completo, mas existem vários exemplos em que as estimativas indicam que mais de 50% das espécies desaparecem no mesmo nível estratigráfico. 6Os limites entre níveis estratigráficos são frequentemente identificados com base em desaparecimentos coordenados. O maior exemplo disso é o desaparecimento de quase metade das famílias (ver Figura 3) e cerca de 95% de todas as espécies no topo do Paleozoico. Dinossauros e muitos outros grupos de répteis e invertebrados marinhos desaparecem do registro no topo do Mesozóico. Outros exemplos de desaparecimentos coordenados em larga escala ocorrem na parte superior do ordoviciano, perto do topo do Devoniano e do topo do Triássico. O desaparecimento coordenado é um padrão de subtração. Não foram relatadas tendências sustentadas neste padrão”.(Gibson, 1996)
Figura 3 - Padrão estratigráfico do número de famílias de invertebrados marinhos representados pelos fósseis em cada nível estratigráfico. (Após Sepkoski 1993, ver Nota 6.)
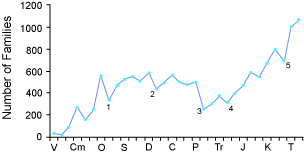
Nível Estratigráfico: V = Vendian; Cm = Cambriano; O = ordovicêutico; S = Siluriano; D = Devoniano; C = Carbonífero; P = Permiano; Tr = Triássico; J = Jurássico; K = Cretáceo; T = Terciário. Pontos marcados como 1-5 representam as cinco maiores "extinções em massa"
A segregação estratigráfica espontânea de corpos e sedimentos comuns, explica uma certa alternância entre presença de muitos fósseis de uma categoria seguido de falta dela (o que é interpretado como extinção em massa) .
O PMS gera muita necessidade de deduções e modelos modelos matemáticos, simulações macroevolutivas em computadores, entre inumeráveis justificativas, que como esforços , como se referiu o Dr Tamborini em relação a paleontologia idiográfica, "são científicos em um sentido puramente descritivo, mas são completamente inúteis para investigações biológicas. Finalmente” (Tamborini, 2015).
Alegações que em 90% dos 544 milhões de anos totais do Fanerozóico, as bio-modificações se estagnaram, repetindo na coluna geológica as mesmas espécies, e só recentemente nas camadas mais atuais (superiores) é que houve biodiversificação em larga escala (radiação) (Gould, 1981) dão um exemplo de descrições científicas terem maior peso do que os dados em si. Além destas justificativas em relação a PMS, temos gigantesca lista de ad hocs justificadores de claros anacronismos geocronológicos como por exemplo, tecidos moles e suposta inesperada preservação de proteínas de animais fossilizados, datados entre 60-120 Ma; onde os autores em vez de questionarem as intocáveis datações absolutas, resolvem buscar formas de justificar incriveis preservações de frágeis proteínas, outro exemplo vem da propria radiometria datacional , com a presença inesperada de carbono 14 datável em diamantes “incontamináveis” de 300 Ma (Baumgardner, 2005);, e quase uma centena de perspectivas e métodos datacionais , que são esquecidos, pela preferência daquilo que combina com o tempo do paradigma geocronológico vigente. Ver TABELA 2.
Figura 4 - Fluxo das interpretações geocronológicas quanto ao PMS, tecidos moles, carbono 14 , onde a geocronologia assumida obriga pesquisadores a fabricarem justificativas ad hoc a direita do que interpretarem o fato em si .

Calcula-se que C14 numa amostra tenha desaparecido totalmente após 100-250 mil anos , já que sua meia vida é de apenas 5730 anos , fato que leva muitos ao dogma de que não se testa coisas supostamente de Ma com C14. No entanto, desafiando esta convenção, foram feitos milhares de testes da equipe do geofísico Dr John Baumgardner no çlaboratorio de Los Álamos, Texas, os quais demonstraram que rochas de 300-500 milhões de anos continham quantificações de teor datável de C14; diante de ad hocs justificando tal anomalia , os críticos recorreram a contaminação, então eles testaram até mesmo em diamantes “incontamináveis” ( a possibilidade de contaminação em diamantes chega a ser desprezível) dando semelhantes resultados e confrontando a geocronologia convencional. (Baumgardner, 2005)
Dr. Tom Kemp, curador das coleções zoológicas do Museu de História Natural da Universidade de Oxford, por exemplo, fez a seguinte admissão: “como é agora bem conhecido, a maioria das espécies fósseis aparecem instantaneamente no registro fóssil, persistem por alguns milhões de anos praticamente inalterados, e apenas desaparecem abruptamente” (Kemp, 1985, p. 67).
Darwin sugeriu que os seres vivos surgiam por evolução gradativa, e esperava que um dia o registro fóssil confirmasse sua predição, porém não foi o que ocorreu: Darwin estava errado. Dia após dia, os inúmeros fósseis escavados por todo o mundo têm refutado a hipótese de mudança gradual em camadas sobrepostas supostamente cronológicas. Ao contrário do que Darwin esperava, os dados recentes revelam padrões de aparecimento súbito (explosões) seguido por longos períodos de pouca mudança.
O famoso paleontólogo evolucionista Niles Eldredge admitiu na revista New Scientist que:
"os paleontólogos desde Darwin têm feito buscas (em grande parte em vão) de sequências de séries graduadas de fósseis que se destacam como exemplos do tipo de transformação global de espécies que Darwin imaginou como o produto natural do processo evolutivo. Poucos viram qualquer razão para duvidar − embora seja um fato surpreendente que [...] a maioria das espécies permanecem reconhecidamente as mesmas, praticamente inalteradas ao longo de sua ocorrência em estratos geológicos de várias idades" (Eldredge, 1986, p.55).
A ausência de fósseis intermediários é demasiado óbvia para os evolucionistas encobrirem por mais tempo. O biólogo evolutivo Dr. David Woodruff da Universidade da Califórnia expressou na revista Science o desapontamento de evolucionistas relativo à ausência no registro fóssil de formas transicionais: "as espécies fósseis permanecem inalteradas durante a maior parte de sua história e o registro não consegue conter um único exemplo de uma transição significativa." (Woodruff, 1980, p.716).
Portanto, diante de tantas admissões, finalmente concordamos com a bióloga evolucionista Lynn Margulis quando diz que:
“não há nenhum gradualismo no registro fóssil. [...] O ‘equilíbrio pontuado’ foi inventado para descrever a descontinuidade no surgimento de novas espécies. [...] Os críticos, [incluindo os críticos SRABUCS], estão certos nas críticas que fazem. [...] Os biólogos evolucionistas acreditam que o padrão evolucionário é uma árvore. Não é!” (Teresi, 2011).
Exemplos de especiação rápida
Existem diversos relatos de surgimento de novas “espécies” em períodos que variam de dezenas há milhares de anos. Segundo essas informações, a especiação é um fenômeno que não necessita de milhões de anos para acontecer. O biólogo norte-americano Dr. James Gibson, Diretor do Instituto de Pesquisa em Geociência (GRI, na sigla em inglês), instituição afiliada à Universidade de Andrews (EUA), mantida pela IASD, descreveu um exemplo de especiação em tempo real: “uma nova espécie de copépode [crustáceos] formou-se no Mar Salton no sul da Califórnia em menos de 30 anos.” (Johnson, 1953; Gibson, 2002).
Estudos diversos têm relatado que apenas entre 10 a 36 anos as populações diferentes de lagartos sofreram alterações morfológicas significativas o suficiente para serem consideradas novas “espécies” (Morrell, 1997;Herrel et al., 2008). Outros exemplos conhecidos de especiação em tempo real incluem bactérias (Shikano et al., 1990), moscas (Huey et al., 2000), tentilhões (Grant e Grant, 2006), rãs (Hoskin et al., 2005), escaravelhos (Halliburton e Gall, 1981) e plantas (Groves e Groves, 1880; Foucaud, 1897; Marchant, 1963).
Dr. Gibson forneceu ainda exemplos de especiação em tempo histórico-arqueológico: “uma população de macacos verdes viveu na ilha de St Kitts no Caribe por menos de 100 anos, mas desenvolveu aspectos morfológicos equivalentes a uma nova espécie.” (Ashton et al., 1979; Gibson, 2002). “O Havaí não tinha bananeiras até cerca de 1000 anos atrás, no entanto há mariposas havaianas nativas que só se alimentam de bananeiras. Estas novas espécies surgiram em menos de 1000 anos.” (Zimmerman, 1960; Gibson, 2002). Diante desses exemplos, Dr. Gibson conclui: “a capacidade de mudanças rápidas está confirmada tanto por experimentação como por observação da natureza” (Gibson, 2002).
TESTEMUNHO ARQUEOLÓGICO
Mais de uma centena de relatos arqueológicos apresentam cenas apavorantes do que ocorreu recentemente na terra . A maioria destes relatos não apresentam aspectos simbólicos, míticos, mas demonstram ser uma descrição literal de um fato dando detalhes as vezes técnicos e matemáticos. Um relato de um livro sagrado dos Maias , o "Chilam Balaam", parece descrever cenas do que poderíamos pensar em termos de consequências de impactos de asteroides na terra, da seguinte forma:
"Ah Mucencab veio e obscureceu a face dos Céus... a Terra começou a despertar. Ninguém sabia o que ia acontecer. De repente, fogos subterrâneos irromperam subindo Firmamento adentro e choveu fogo do alto, e despencaram cinzas, e pedras e árvores foram lançadas para baixo, e madeiras e pedras se despedaçaram umas contra as outras. Então o Céu foi agarrado e afastado à força. A face do Céu foi fustigada de um lado para o outro e atirada para trás... as pessoas foram todas esmigalhadas; seus corações pararam enquanto elas ainda viviam. Então elas foram enterradas nas areias, no mar. Numa enorme e repentina torrente de água, a Grande Serpente foi arrebatada do Céu. O Firmamento caiu e a Terra afundou quando os quatro deuses, os Bacabs, apresentaram quem trouxe a destruição do mundo." . Os Incas disseram : Incas "Um dia, um grupo de estrangeiros chegou para avisar que os orgulhosos habitantes daquela cidade seriam destruídos pelo terremoto, pela inundação e pelo fogo. A maioria deles riu dos estrangeiros. Contudo, alguns sacerdotes da cidade ouviram o aviso e foram viver em lugares afastados no topo das montanhas. Algum tempo depois, uma nuvem vermelha apareceu no horizonte.Ela cresceu e cobriu toda a área, e sua vermelhidão era tão forte que iluminava o céu noturno. De repente, com um brilho e um estrondo, um terremoto destruiu muitos edifícios da cidade, e uma chuva vermelha começou a cair.Outros terremotos e mais chuva vieram em sequência, e uma inundação logo cobriu toda a cidade arruinada" Outro relato dos maias esse retirado do Popul Vuh: "Então as águas foram agitadas pela vontade de Hurakán, e uma grande inundação desabou sobre as cabeças dessas criaturas... Elas foram tragadas e uma nebulosidade resinosa desceu do céu... a face da Terra ficou sombria e uma pesada chuva que tudo escurecia começou a cair — chuva de dia e chuva de noite... Ouviu-se um grande barulho acima de suas cabeças, como se produzido pelo fogo. Então se viram homens correndo, empurrando-se, cheios de desespero; quiseram subir em suas casas, e as casas, desmoronando, caíram por terra; quiseram trepar nas árvores, e as árvores fizeram chacoalharam à sua frente"
Conclusão
Padrões morfológicos em torno do táxon gênero (PMTG) se identificam com fósseis ancestrais básicos sepultados num catastrofismo recente (SRABUC), pois este modelo respeita o fato da especiação rápida observada, que exigiria uma variabilidade proporcional muito grande no registro fóssil, caso representasse a evolução em milhões de anos (PMS), e isto além de não existir tal variação taxonômica, ainda se confirma com estase morfológica, repetição das mesmas espécies e 4229 gêneros de fósseis vivos, quadro este que reflete um sepultamento repentino dos seres vivos do planeta e não uma suposta história evolutiva da vida de Ma. Relatos de especiação na história e arqueologia, capazes de justificar toda biodiversidade em pouco tempo, nos mostram que deveríamos ter muito mais variabilidade no registro fóssil do que há, o que, dado o fato da evolução ser verificada, podemos certificar que não houve tempo para que ela atuasse. A explosão cambriana, a ausência de variabilidade amostral fóssil (estase morfológica fóssil e pobreza taxonômica) e falta de radiação especiativa no registro fóssil (diversificação) até o pleistoceno, nos conta uma história de um período de: 1) surgimento rápido e pronto de formas de vida , 2) permanência morfológica com alto número de espécies em ambiente estável (repetição fóssil sem pressões ambientais evolutivas e adaptativas, e sem grandes catástrofes intercalando tempo), 3) desastre soterrador de população viva evidenciada pelas repetições de mesmas espécies fósseis (o que descaracteriza a pontualidade), 4) presença de diversas espécies diferentes unidas no registro fóssil, fósseis de imensos vertebrados completos continentais(o que caracteriza desastre de grande magnitude e altas taxas de sedimentação), e 5) mudança drástica no ambiente gerando a radiação das espécies (diversificação) nas camadas recentes de forma amostral (fósseis recentes) e nas milhões de espécies na biodiversidade atual. “Enquanto há 200 anos, os naturalistas achavam que havia talvez cinco ou dez mil espécies na Terra, estimativas atuais... coloque o valor em dez ou 20 milhões. Costuma-se supor que a vida nunca tenha sido mais diversificada do que é hoje”. (Benton et al, 2007) Estes pontos levantados, questionam a geocronologia , e não buscam , como muitos de artigos científicos o fazem, justificar com ad hocs as anomalias anacrônicas do que se esperaria encontrar nos fatos paleontológicos, mas destacam inteerpretações mais independentes da geocronologia convencional, sugerindo assim amplo questionamento das datações de alta escala de tempo , chamadas de “absolutas” e em especial, da hipótese uniformitarianista de que nada acelerou o decaimento radioativo envelhecendo rochas durante os supostos bilhões de anos aferidos, que ainda imperam por décadas sobre o pensamento dinâmico e científico moderno, mesmo possuindo talvez centenas de métodos e aperfeiçoamentos para acelerar decaimento radioativo e digamos assim, envelhecer as rochas com voltagens e ambientes que representam uma irrisória comparação com o que ocorreu quando bólidos imensos cairam sobre a terra, onde aproximadamente 99,8% foram apagados pela dinâmica tectônica, marítma , erosiva e sedimentar da terra.
A estase morfológica no registro fóssil só pode representar o sepultamento de uma população em um só momento , qualquer interpretação que avançar um milímetro sequer desta realidade, contrariará tudo que conhecemos sobre evolução e plasticidade adaptativa das espécies ao longo do tempo e a hipótese de uma chuva de asteroides parece ser suficiente para poder não somente explicar o PMS, mas quase todas as anomalias geoquímicas e geocronológicas da terra. Milhares de estudos poderão ser desenvolvidos a partir destas poucas reflexões introdutórias que fizemos aqui.
REFERÊNCIAS
Goodings, CR & Brookfield, ME, 1992.Movimentos transcorrentes proterozóicos ao longo do linfonodo Kapuskasing (Província Superior, Canadá) e sua relação com as estruturas circundantes. Earth-Science Reviews, 32: 147-185.
Bleeker, W. e Pilkington, M., 2004.O Arco Nastapoka de 450 km de diâmetro: a cicatriz de impacto preservada mais antiga e maior da Terra? Programa com resumos - Associação Geológica do Canadá; Associação Mineralógica do Canadá: Reunião Anual Conjunta, 2004, Vol. 29, pp. 344.
Beals, CS, 1968. Sobre a possibilidade de uma origem catastrófica para o grande arco do leste da baía de Hudson. Em: Beals, CS (Ed.), Pp. 985-999. Ciência, História e Hudson Bay , Vol. 2, Ministério das Minas e Recursos da Energia, Ottawa.
1. Genesis 1:1; 7:11,12
2. Chilam Balam : (Chumayel). Barcelona: Linkgua Ediciones. 2008. ISBN 9788498971439. OCLC 502270722
3. Aaron M (2014). Baraminological Analysis of the Caseidae (Synapsida: Pelycosauria). Journal of Creation Theology and Science Series B: Life Sciences. 4:19-22.
4. Albrecht C, Wilke T (2008). Ancient Lake Ohrid: biodiversity and evolution. Hydrobiologia. 615(1):103-140.
5. Alisson E (2013). Arqueologia ajudará a desvendar origem da biodiversidade amazônica. Agência FAPESP. Disponível em: http://agencia.fapesp.br/arqueologia_ajudara_a_desvendar_origem_da_biodiversidade_amazonica/16937/
6. Ariza LM (2007). Evolution in a Petri Dish. Scientific American. 297(6):36. Disponível em: http://www.scientificamerican.com/article/evolution-in-a-petri-dish/
7. Ashton EH, Flinn RM, Griffiths RK, Moore WJ (1979). The results of geographic isolation on the teeth and skull of the Green monkey (Cercopithecus aethiopssabaeus) in St. Kitts – a multivariate retrospect. Journal of Zoology. 188(4):533-555.
8. Aw Kinderewitsch e Li Kitscha: Polewaja suschtschnost jadernoi fisiki - campo de Física Nuclear, Kiev: EKMO, 2003, páginas 263 -30
Basener, William F.; Sanford, John C. (7 de novembro de 2017). «The fundamental theorem of natural selection with mutations». Journal of Mathematical Biology(em inglês): 1–34. ISSN 0303-6812. doi:10.1007/s00285-017-1190-x
Barton, PJ, Grieve, RAF, Morgan, JV, Surendra, AT, Vermeesch, PM, Christeson, GL, Gulick, SPS e Warner, MR, Imagens sísmicas da folha de derretimento de impacto Chicxulub e comparação com a estrutura de Sudbury; em: Gibson, RL e Reimold, WU (Eds.), grandes impactos de meteoritos e evolução planetária IV , papel especial GSA 465, pp. 103-113, 2010.
9. Baumgardner, Jonh (2005). «Radioisotopes and the Age of the Earth, Volume II». Institute for Creation Research. Consultado em 10 de setembro de 2017
10. Behe, M. June 1997). «Darwinism and design». Trends in Ecology & Evolution. 12 (6). 229 páginas. ISSN 0169-5347. PMID 2. (J1238050
11. Behe, Michael J. (February 2009). «Waiting longer for two mutations». Genetics. 181 (2): 819–820; author reply 821–822. ISSN 0016-6731. PMID 19189948. doi:10.1534/genetics.108.098905
12. Bell G (2013). Evolutionary rescue and the limits of adaptation. Philos Trans R Soc Lond B Biol Sci. 368(1610):20120080.
13. Benedict, John C.; Smith, Selena Y.; Specht, Chelsea D.; Collinson, Margaret E.; Leong-Škorničková, Jana; Parkinson, Dilworth Y.; Marone, Federica (2016). «Species diversity driven by morphological and ecological disparity: a case study of comparative seed morphology and anatomy across a large monocot order». AoB PLANTS. 8. ISSN 2041-2851. PMID 27594701
14. Benton, Michael J.; Emerson, Brent C. (1 de janeiro de 2007). «How Did Life Become so Diverse? the Dynamics of Diversification According to the Fossil Record and Molecular Phylogenetics». Palaeontology (em inglês). 50 (1): 23–40. ISSN 1475-4983. doi:10.1111/j.1475-4983.2006.00612.x
15. Berthault, G. : “Orogenesis: Cause of sedimentary formations” – Kazan Golovkinsy Stratigraphic Meeting, 2014, pp.19-20
16. Berthault, G. : “Orogenesis: cause of sedimentary formations” – “Open Journal of Geology“ ISSN 2161-7570.Vol 3, Number 28, April 2013.
17. Berthault G. : “Towards a Refoundation of Historical Geology” – “Georesources” 1(12) 2012, p.38, 39
18. Berthault, G., Lalomov, A. V. and Tugarova, M. A. : “Reconstruction of paleolithodynamic formation conditions of Cambrian-Ordovician sandstones in the Northwestern Russian platform” – “Lithology and Mineral Resources, 2011, Volume 46, Number 1, 60-70” (Springer Publishing site)
19. Berthault, G., Veksler A.B., Donenberg V.M. , Lalomov A. : “RESEARCH on EROSION OF CONSOLIDATED and semi-consolidated SOILS BY HIGH SPEED WATER FLOW” Izvestia.VNIIG., 2010, Vol. 257, pp.10-22. – (Russian original.)
20. Berthault, G : “Sedimentological Interpretation of the Tonto Group Stratigraphy (Grand Canyon Colorado River)” , Lithology and Mineral Resources 2004, Vol. 39, No 5. October 2004.
21. Berthault G., “Analysis of Main Principles of Stratigraphy on the Basis of Experimental Data”, Litol.Polezn.Iskop.2002, vol 37, no.5,pp 509-515 (Lithology and Mineral resources 2002 (fac-similé) (Engl.Transl.), vol.37, no.5, pp442-446), Journal of the Academy of Sciences of Russia.
22. Berthault, G., “Sedimentation of a Heterogranular Mixture. Experimental Lamination in Still and Running Water”, Compte rendu de l’Académie des Sciences 1988, vol. 306, Serie II, pp. 717–724.
23. Berthault, G., “Sedimentologie: Expériences sur la lamination des sédiments par granoclassement périodique postérieur au dépôt. Contribution a l’explication de la lamination dans nombre de sédiments et de roches sédimentaires”., Compte rendu de l’Académie des Sciences de Paris 1986 , vol. 303, Ser., 2, no. 17, pp. 1569-1574.
24. Bhullar BS (2017). Evolution: Catastrophe triggers diversification. Nature. 542 (7641):304-305.
25. Bosch, Fritz (1999). «Setting a Cosmic Clock with Highly Charged Ions». Physica Scripta (em inglês). T80 (A). ISSN 1402-4896. doi:10.1238/physica.topical.080a00028
26. Brand LR, Tang T (1991). Fossil vertebrate footprints in the Coconino Sandstone (Permian) of northern Arizona: Evidence for underwater origin. Geology. 19(12):1201-1204.
27. Brawn, Dan (2013). «The Origin of Earth's Radioactivity». Consultado em 24 de setembro de 2017
28. Bunch, Ted E.; Hermes, Robert E.; Moore, Andrew M. T.; Kennett, Douglas J.; Weaver, James C.; Wittke, James H.; DeCarli, Paul S.; Bischoff, James L.; Hillman, Gordon C. (10 de julho de 2012). «Very high-temperature impact melt products as evidence for cosmic airbursts and impacts 12,900 years ago». Proceedings of the National Academy of Sciences of the United States of America. 109 (28): E1903–1912. ISSN 1091-6490. PMID 22711809. doi:10.1073/pnas.1204453109
29. Carroll, R. L. (1992). «The Primary Radiation of Terrestrial Vertebrates». Annual Review of Earth and Planetary Sciences. 20 (1): 45–84. doi:10.1146/annurev.ea.20.050192.000401
30. Catalogue of Life (2016). Species 2000 & ITIS Catalogue of Life, 2016 Annual Checklist. Disponível em: http://www.catalogueoflife.org/col/info/ac
31. Cavanaugh DP, Sternberg RV (2004). Analysis of morphological groupings using ANOPA, a pattern recognition and multivariate statistical method: A case study involving centrarchid fishes. Journal of Biological Systems. 12(2):137–67.
32. Cavanaugh DP, Wood TC (2002). A Baraminological Analysis of the Tribe Heliantheaesensulato (Asteraceae) Using Analysis of Pattern (ANOPA). Occas. Papers of the BSG. 17(1):1-11.
33. Chadwick A, Spencer L (2006). Turner Preliminary depositional model for an Upper Cretaceous Edmontosaurusbonebed. Journal of Vertebrate paleontology. 26:49A.
34. Crabtree GR (2013a). Our fragile intellect. Part I. Trends in Genetics. 29(1):1-3.
35. Crabtree GR (2013b). Our fragile intellect. Part II. Trends in Genetics. 29(1):3-5.
36. Crow JF (1997). The high spontaneous mutation rate: is it a health risk? Proc Natl AcadSci U S A. 94(16):8380-6.
37. Darwin C (1861). On the Origin of Species. 3. Ed. London: Murray. capítulo 14.
38. de Paula MO (2009). Podem os criacionistas aceitar a origem de novas espécies? Earth History Research Center. Disponível em: http://origins.swau.edu/papers/evol/marcia1/defaultp.html
39. de Queiroz K (2005). Ernst Mayr and the modern concept of species. Proc Natl AcadSci U S A. 102 Suppl 1:6600-7.
40. Dilly, R., Berthault, G.: “Orogenesis: Cause of sedimentary formations” – The Russian Academy of Sciences scientific Council on Lithology and Minerals in Sedimentary Formations – VIII All-Russian Lithological Meeting (Moscow, 27-30 October 2015), Tome II, pp. 162-164
41. Dolgin E (2009). Human mutation rate revealed. Nature News (27 Ago. 2009). Disponível em:http://www.nature.com/news/2009/090827/full/news.2009.864.html
42. Donnelley, Strachan (July 2002). «Natural responsibilities. Philosophy, biology, and ethics in Ernst Mayr and Hans Jonas». The Hastings Center Report. 32(4): 36–43. ISSN 0093-0334. PMID 12362522
43. Eakin CM (2014). Lamarck was partially right—and that is good for corals. Science. 344(6186):798-9.
44. Eldredge N, Stanley SM (1984). Living Fossils: Introduction to the Casebook. New York/Berlin: Springer-Verlag pp. 266-270.
45. Eldredge N (1986). Progress in Evolution? New Scientist. 110:54-57.
46. Eyre-Walker A, Keightley PD (1999). High genomic deleterious mutation rates in hominids. Nature. 397(6717):344-7.
47. Ezard, Thomas H. G.; Quental, Tiago B.; Benton, Michael J. (5 de abril de 2016). «The challenges to inferring the regulators of biodiversity in deep time». Phil. Trans. R. Soc. B(em inglês). 371 (1691). 20150216 páginas. ISSN 0962-8436. PMID 26977058. doi:10.1098/rstb.2015.0216
48. Grant, B. Rosemary; Grant, Peter R. (3 de março de 2017). «Watching speciation in action». Science (New York, N.Y.). 355 (6328): 910–911. ISSN 1095-9203. PMID 28254901. doi:10.1126/science.aam6411.
49. Folha de S. Paulo (2006). Pássaros exibem seleção natural em tempo real. Folha de São Paulo (14/07/2006). Disponível em: http://www1.folha.uol.com.br/folha/ciencia/ult306u14848.shtml
50. Fondon JW, Garner HR (2004). Molecular origins of rapid and continuous morphological evolution. Proc Natl AcadSci U S A. 101(52):18058-63.
51. Foote, Mike (1993/04). «Discordance and concordance between morphological and taxonomic diversity». Paleobiology. 19 (2): 185–204. ISSN 0094-8373. doi:10.1017/S0094837300015864
52. Foucaud (1897). Un Spartinainédit. Ann SocSci Nat Char Inf. 32:220–222.
53. Frair W (2000). Baraminology: Classification of Created Organisms. CRSQ Quarterly. 37(2):82-91.
54. Freeman S, Herron JC (2009). Análise evolutiva. 4. ed. orto Alegre: Artmed. 848p.
55. Fu W, et al (2013). Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants. Nature. 493(7431):216-20.
56. Futuyma, Douglas J. (July 2010). «Evolutionary constraint and ecological consequences». Evolution; International Journal of Organic Evolution. 64(7): 1865–1884. ISSN 1558-5646. PMID 20659157. doi:10.1111/j.1558-5646.2010.00960.x
57. Furness, Andrew I.; Lee, Kevin; Reznick, David N. (June 2015). «Adaptation in a variable environment: Phenotypic plasticity and bet-hedging during egg diapause and hatching in an annual killifish». Evolution; International Journal of Organic Evolution. 69 (6): 1461–1475. ISSN 1558-5646. PMID 25908306. doi:10.1111/evo.12669
58. Gentry, R.V. 1970. "Giant Radioactive Halos: Indicators of Unknown Alpha-Radioactivity?" Science 169, 670.
59. Gentry, R.V. 1971. "Radiohalos: Some Unique Pb Isotope Ratios and Unknown Alpha Radioactivity." Science 173, 727.
60. Gentry, R.V. 1973. "Radioactive Halos." Annual Review of Nuclear Science 23, 347. Acessado em 24/09/2017 em http://www.robertvgentry.com/
61. Gentry, R.V. 1974. "Radiohalos in Radiochronological and Cosmological Perspective." Science 184, 62. Acessado em 24/09/2017 em http://www.robertvgentry.com/
62. Gentry, R.V. 1977. "Mystery of the Radiohalos." Research Communications NETWORK, Breakthrough Report, February 10, 1977. Acessado em 24/09/2017 em http://www.robertvgentry.com/
63. Gentry, R.V. 1978a. "Are Any Unusual Radiohalos Evidence for SHE?" International Symposium on Superheavy Elements, Lubbock, Texas. New York: Pergamon Press.
64. Gentry, R.V. 1978b. "Implications on Unknown Radioactivity of Giant and Dwarf Haloes in Scandinavian Rocks." Nature 274, 457.
65. Gentry, R.V. 1978c. "Reinvestigation of the α Activity of Conway Granite." Nature 273, 217. PDF
66. Gentry, R.V. 1979. "Time: Measured Responses." EOS Transactions of the American Geophysical Union 60, 474. Acessado em 24/09/2017 em http://www.robertvgentry.com/
67. Gentry, R.V. 1980. "Polonium Halos." EOS Transactions of the American Geophysical Union 61, 514.
68. Gentry, R.V. et al. 1973. "Ion Microprobe Confirmation of Pb Isotope Ratios and Search for Isomer Precursors in Polonium Radiohalos." Nature 244, 282.
69. Gentry, R.V. et al. 1974. "'Spectacle' Array of Po-210 Halo Radiocentres in Biotite: A Nuclear Geophysical Enigma." Acessado em 24/09/2017 em http://www.robertvgentry.com/
70. Gentry, R.V. et al. 1976a. "Radiohalos and Coalified Wood: New Evidence Relating to the Time of Uranium Introduction and Coalification." Science 194, 315 . Acessado em 24/09/2017 em http://www.robertvgentry.com/
71. Ghalambor, Cameron K.; Hoke, Kim L.; Ruell, Emily W.; Fischer, Eva K.; Reznick, David N.; Hughes, Kimberly A. (17 de setembro de 2015). «Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature». Nature (em inglês). 525 (7569): 372–375. doi:10.1038/nature15256
72. Gibson LJ (2002). A similaridade biológica e a teoria da evolução. In: 4º Encontro Nacional de Criacionistas, 24 a 28 de janeiro de 2002, São Paulo. Anais... São Paulo: Centro Universitário Adventista, campus São Paulo.
73. Gibson, L. J. --- Fossil Patterns: A Classification and Evaluation». www.grisda.org. Consultado em 10 de setembro de 2017
74. Gorodezki V, 2005. Deactivation unit for radioactive materials, comprises capacitor with curved mantle surfaces, connected to controllable generator DE 10354251 A1.Acessado em 24/09/2017 em http://www.google.com.na/patents/DE10354251A1?cl=en&hl=pt-BR
75. Gould SJ (1981). Evolution as Fact and Theory, Discover magazine 1981; 2(5):34-37; Reimpresso com permissãoem: Gould SJ. Hen's Teeth and Horse's Toes. New York: W. W. Norton & Company, 1994, pp. 253-262.
76. Grant PR, Grant BR (2006). Evolution of Character Displacement in Darwin's Finches. Science. 313(5784):224-6.
77. Groves H, Groves J (1880). Spartinatownsendiinobis. Rep Bot SocExch Club Br Id. 1:37.
78. Donald R. Lowe, Gary R. Byerly, Frank T. Kyte, Alexander Shukolyukov, Frank Asaro e Alexandra Krull. Astrobiologia. Julho de 2004, 3 (1): 7-48. https://doi.org/10.1089/153110703321632408
79. Halliburton R, Gall GAE (1981). Disruptive selection and assortative mating in Triboliumcastaneum. Evolution. 35:829-843.
80. Herrel A, et al (2008). Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource. Proc Natl AcadSci U S A. 105(12):4792-5.
81. Hesslow, L.; Embréus, O.; Stahl, A.; DuBois, T. C.; Papp, G.; Newton, S. L.; Fülöp, T. (23 de junho de 2017). «Effect of Partially Screened Nuclei on Fast-Electron Dynamics». Physical Review Letters. 118 (25). 255001 páginas. ISSN 1079-7114. PMID 28696736. doi:10.1103/PhysRevLett.118.255001
82. Horner JR, Goodwin MB (2009). Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS One. 4(10):e7626.
83. Hoskin CJ1, Higgie M, McDonald KR, Moritz C (2005). Reinforcement drives rapid allopatric speciation. Nature. 437(7063):1353-6.
84. Huey RB, et al (2000). Rapid Evolution of a Geographic Cline in Size in an Introduced Fly. Science. 287(5451):308-309.
85. I Have A List"». Eye on the ICR (em inglês). 31 de julho de 2011
86. Izumi, Y.; Tomita, H.; Nakayama, Y.; Hayashi, S.; Morishima, K.; Isobe, M.; Cheon, M. S.; Ogawa, K.; Nishitani, T. (November 2016). «Development of fast neutron pinhole camera using nuclear emulsion for neutron emission profile measurement in KSTAR». The Review of Scientific Instruments. 87 (11): 11D840. ISSN 1089-7623. PMID 27910454. doi:10.1063/1.4963866
87. Jerlström P (2000). Is the evolutionary tree turning into a creationist orchard? Journal of Creation. 14(2):11–13.
88. Johnson MW (1953). The copepod Cyclops dimorphus Kiefer from the Salton Sea. American Midland Naturalist. 49:188-192.
89. Julien, P.Y., Lan, Y., and Berthault, G., “Experiments on Stratification of Heterogeneous Sand Mixtures”, Bulletin Société Géologique de France, 1993, vol. 164, no. 5, pp. 649–660.
90. Junker R, Scherer S (1996). Evollução: um livro texto crítico. 4. Ed. Brasília, DF: Sociedade Criacionista Brasileira.
91. Kahle H (1999). Evolution: IrrwegmodernerNaturwissenschaft? 4. Ed. Bielefeld: Mindt, Edeltraud. 200p.
92. Keightley PD (2012). Rates and Fitness Consequences of New Mutations in Humans. Genetics. 190(2):295-304.
93. Kemp TS (1985). A Fresh Look at the Fossil Record. New Scientist. 108(1485):66-67.
94. Kennett, James P.; Kennett, Douglas J.; Culleton, Brendan J.; Aura Tortosa, J. Emili; Bischoff, James L.; Bunch, Ted E.; Daniel, I. Randolph; Erlandson, Jon M.; Ferraro, David (11 de agosto de 2015). «Bayesian chronological analyses consistent with synchronous age of 12,835-12,735 Cal B.P. for Younger Dryas boundary on four continents». Proceedings of the National Academy of Sciences of the United States of America. 112 (32): E4344–4353. ISSN 1091-6490. PMID 26216981. doi:10.1073/pnas.1507146112
95. Kennett, James P.; Kennett, Douglas J.; Culleton, Brendan J.; Aura Tortosa, J. Emili; Bischoff, James L.; Bunch, Ted E.; Daniel, I. Randolph; Erlandson, Jon M.; Ferraro, David (11 de agosto de 2015). «Bayesian chronological analyses consistent with synchronous age of 12,835-12,735 Cal B.P. for Younger Dryas boundary on four continents». Proceedings of the National Academy of Sciences of the United States of America. 112 (32): E4344–4353. ISSN 1091-6490. PMID 26216981. doi:10.1073/pnas.1507146112
96. Kuhn, Joseph A. (2012-1). «Dissecting Darwinism». Proceedings (Baylor University. Medical Center). 25 (1): 41–47. ISSN 0899-8280. PMID 22275784
97. Lacy, Robert C. (1 de agosto de 1987). «Loss of Genetic Diversity from Managed Populations: Interacting Effects of Drift, Mutation, Immigration, Selection, and Population Subdivision». Conservation Biology (em inglês). 1 (2): 143–158. ISSN 1523-1739. doi:10.1111/j.1523-1739.1987.tb00023.x
98. Lalomov, A., Berthault G., Tugarova, M., Isotov V., Sitdikova L.: “Reconstruction of sedimentary conditions of Middle Permian Kama-Ural basin studied by N.A.Golovkinsky” – Kazan Golovkinsy Stratigraphic Meeting, 2014, pp.53-54
99. Lalomov, A. : “Reconstruction of Paleohydrodynamic Conditions during the Formation of Upper Jurassic Conglomerates of the Crimean Peninsula”, Lithology and Mineral Resources, 2007, Vol. 42, No. 3, pp. 268–280
100. Lavoué, Sébastien; Miya, Masaki; Arnegard, Matthew E.; McIntyre, Peter B.; Mamonekene, Victor; Nishida, Mutsumi (7 de abril de 2011). «Remarkable morphological stasis in an extant vertebrate despite tens of millions of years of divergence». Proceedings of the Royal Society B: Biological Sciences. 278 (1708): 1003–1008. ISSN 0962-8452. PMID 20880884.
101. Lee, Kanani K. M.; Steinle-Neumann, Gerd (30 de março de 2008). «Ab-initio study of the effects of pressure and chemistry on the electron-capture radioactive decay constants of 7Be, 22Na and 40K». Earth and Planetary Science Letters. 267 (3): 628–636. doi:10.1016/j.epsl.2007.12.014
102. Levinton JS, Chris MS (1980). A Critique of the Punctuated Equilibria Model and Implications for the Detection of Speciation in the Fossil Record. Syst Zool. 29(2):130–142.
103. Lightner JK (2016). Variation after the Flood. Creation Matters. 21(2):6-7.
104. Locey KJ, Lennon JT (2016). Scaling laws predict global microbial diversity. Proc Natl AcadSci U S A. 113(21):5970-5.
105. Lönnig, Wolf-Ekkehard (2004). «Dynamic genomes, morphological stasis, and the origin of irreducible complexity». Max-Planck-Institut for Plant Breeding Research. Consultado em 9 de setembro de 2017
106. Lopes RJ (2015). Afobação faz cientistas classificarem fósseis de símios como hominídeos. Folha de S. Paulo. Disponível em: http://m.folha.uol.com.br/ciencia/2015/09/1676679-no-afa-cientistas-tem-classificado-fosseis-de-simios-como-hominideos.shtml?mobile
107. Lynch M (2010). Rate, molecular spectrum, and consequences of human mutation.Proc Natl AcadSci U S A. 107(3):961-8.
108. Marchant CJ (1963). Corrected chromosome numbers for Spartina x townsendii and its parent species. Nature. 199:929.
109. Marsh FL (1941). Fundamental Biology. Lincoln, NE: Published by the Author.
110. Marsh FL (1978). Variation And Fixity Among Living Things. A New Biological Principle.CRS Quarterly. 15(2):115-118.
111. Martens K (1997). Speciation in ancient lakes. Trends EcolEvol. 12(5):177-82.
112. Mendonça de Souza, S. M. F., Rodrigues-Carvalho, C., Silva, H. P., & Locks, M. (2006). Revisitando a discussão sobre o Quaternário de Lagoa Santa e o povoamento das Américas: 160 anos de debates científicos. In H. P. Silva & C. Rodrigues-Carvalho (Eds.), Nossa origem. O povoamento das Américas – visões multidisciplinares (pp. 19–43). Rio de Janeiro: Viera & Lentz.
113. Minoletti, Fabrice; Hermoso, Michaël; Gressier, Vincent (2009). «Separation of sedimentary micron-sized particles for palaeoceanography and calcareous nannoplankton biogeochemistry». Nature Protocols. 4 (1): 14–24. ISSN 1750-2799. PMID 19131952. doi:10.1038/nprot.2008.200
114. Mora C, Tittensor DP, Adl S, Simpson AG, Worm B (2011). How many species are there on Earth and in the ocean? PLoS Biol. 9(8):e1001127.
115. Morrell V (1997). Catching Lizards in the Act of Adapting. Science. 276(5313):682-683.
116. Nachman MW, Crowell SL (2000). Estimate of the Mutation Rate per Nucleotide in Humans. Genetics. 156(1):297-304.
117. Nagalingum NS, et al (2011). Recent Synchronous Radiation of a Living Fossil. Science. 334 (6057): 796-799.
118. NASA Astrobiology Institute». nai.nasa.gov. Consultado em 24 de setembro de 201
119. Prüfer K, et al (2013). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 505:43-49.
120. ReMine WJ (1993). The Biotic Message. St Paul, MN: St Paul Science.
121. Reinhard Junker, Siegfried Scherer: Evolution – Ein kritisches Lehrbuch. Gießen, ISBN 3-921046-10-6
122. Ridley M (2006). Evolução. 3. ed. Porto Alegre: Artmed. 752p.
123. Robinson DA, Cavanaugh DP (1998). A quantitative approach to baraminology with examples from the primates. CRSQ Quarterly. 34:196–208.
124. Robinson DA (1997). A mitochondrial DNA analysis of the testudineapobaramin. Creation Research Society Quarterly. 33:262–72.
125. Romer AS (1966). Vertebrate Paleontology. 3rd Ed. Chicago: University of Chicago Press, pp. 347–396. In: Denton M. Evolution: A Theory in Crisis. Bethesda: Adler & Adler, 1986, p. 202.
126. Sadava D, Heller HC, Orians GH, Purves WK, Hillis DM (2009). Vida: A Ciência da Biologia. 8. Ed., v. 2. Porto Alegre: EditoraArtmed.
127. Sadler, Peter M. (1 de setembro de 1981). «Sediment Accumulation Rates and the Completeness of Stratigraphic Sections». The Journal of Geology. 89 (5): 569–584. ISSN 0022-1376. doi:10.1086/628623
128. Sanford, John & Baumgardner, John & Brewer, Wesley & Gibson, Paul & Remine, Walter. (2008). Using Numerical Simulation to Test the Validity of Neo-Darwinian Theory.
129. Sanford John C. (2014). Genetic Entropy. 4. ed. New York: FMS Publications.
130. Scherer S (1993). Typen des Lebens. Berlin: Pascal.
131. Schwartz JH, Tattersall I (2015). Defining the genus Homo. Science. 349(6251):931-2.
132. Senter P (2010). Using creation science to demonstrate evolution: application of a creationist method for visualizing gaps in the fossil record to a phylogenetic study of coelurosaurian dinosaurs. Journal of Evolutionary Biology. 23:1732–1743.
133. Sepkoski, J. J. (1993). «Ten years in the library: new data confirm paleontological patterns». Paleobiology. 19 (1): 43–51. ISSN 0094-8373. PMID 11538041
134. Sepkoski, David (2013). «Towards "a natural history of data": evolving practices and epistemologies of data in paleontology, 1800-2000». Journal of the History of Biology. 46 (3): 401–444. ISSN 0022-5010. PMID 22956055. doi:10.1007/s10739-012-9336-6
135. Shabalina SA, Ogurtsov AY, Kondrashov VA, Kondrashov AS (2001). Selective constraint in intergenic regions of human and mouse genomes. Trends Genet. 17(7):373-6.
136. Schopf JW. 1992. Patterns of Proterozoic microfossil diversity: an initial, tentative, analysis. In: Schopf and Klein, p 529-552; (b) Walter MR, Grotzinger JP, Schopf JW. 1992. Proterozoic stromatolites. In: Schopf and Klein, p 253-260
137. Shikano S, Luckinbill LS, Kurihara Y (1990). Changes of traits in a bacterial population associated with protozoal predation. Microbial Ecology. 20:75-84.
138. Snelling A (1997). Sedimentation experiments: Nature finally catches up! Journal of Creation. 11(2):125-126.
139. Souza Jr NN (2008). Uma breve história da terra. Brasília, DF: Sociedade Criacionista Brasileira.
140. Spray, John G.; Kelley, Simon P.; Rowley, David B. (12 de março de 1998). «Evidence for a late Triassic multiple impact event on Earth». Nature (em inglês). 392 (6672): 171–173. ISSN 0028-0836. doi:10.1038/32397
141. Teresi D (2011). Discover Interview: Lynn Margulis Says She's Not Controversial, She's Right. Discover Magazine, p. 68.
142. Trivedi BP (2000). Island mice may evolve faster: From one species to six. Genome News Network. Disponível em: http://www.genomenewsnetwork.org/articles/04_00/island_mice.shtml
143. Van Bocxlaer B, Hunt G (2013). Morphological stasis in an ongoing gastropod radiation from Lake Malawi. PNAS. 110(34):13892-13897.
144. Voje KL (2016). Tempo does not correlate with mode in the fossil record. Evolution. 70(12):2678-2689.
145. Xie, Xufei; Chen, Zhongjing; Peng, Xingyu; Yuan, Xi; Zhang, Xing; Gorini, Giuseppe; Cui, Zhiqiang; Du, Tengfei; Hu, Zhimeng (October 2014). «Neutron emission measurement at the HL-2A tokamak device with a liquid scintillation detector». The Review of Scientific Instruments. 85 (10). 103506 páginas. ISSN 1089-7623. PMID 25362392. doi:10.1063/1.4896120
146. Weyrich A, et al (2016). Paternal intergenerational epigenetic response to heat exposure in male Wild guinea pigs. Mol Ecol. 25:1729-1740.
147. Whitmore J (2013a). Temporal Patterns in 'Living Fossils'. In: 4th Annual Research and Scholarship Symposium. Cedarville, OH. Anais... Cedarville, OH: Cedarville University. Disponível em: http://digitalcommons.cedarville.edu/research_scholarship_symposium/2013/poster_presentations/22/
148. Whitmore J (2013b). What about living fossils? Capítulo 12, pp.143-150. In: Ham K (Ed.). The New Answers Book, v. 4. Green Forest: Master Books.
149. Wills, Matthew A.; Briggs, Derek E. G.; Fortey, Richard A. (1994/04). «Disparity as an evolutionary index: a comparison of Cambrian and Recent arthropods». Paleobiology. 20 (2): 93–130. ISSN 0094-8373. doi:10.1017/S009483730001263X
150. Williamson PG (1981). Morphological Stasis and Developmental Constraint: Real Problems for Neo-Darwinism. Nature. 294:214-2015.
151. Wilmer JW, et al (2011). Catastrophic Floods May Pave the Way for Increased Genetic Diversity in Endemic Artesian Spring Snail Populations. PLoS One. 6(12):e28645.
152. Wilson G (2010). Classic Multidimensional Scaling isn’t the Sine Qua Non of Baraminology. Answers in Genesis. Disponível em: https://answersingenesis.org/creation-science/baraminology/classic-multidimensional-scaling-and-baraminology/
153. Wise KP (1989) The Estimation of True Taxonomic Durations from Fossil Occurrence Data; University Microfilms, 1989.
154. Wise KP (1990). Baraminology: a young-earth creation biosystematic method. In: Walsh RE (Ed.). Proceedings of the Second International Conference on Creationism. Volume II, Technical Symposium, pp. 345–360. Creation Science Fellowship.
155. Wise KP (1992). Practical baraminology. Creation ex Nihilo Technical Journal. 6(2): 122–37.
156. Wise KP (2010) Relatório Gmail [8.7 milion x 300.000]. Mensagem recebida por <sodre.siman@gmail.com> em 13 out. 2015
157. Wittke, James H.; Weaver, James C.; Bunch, Ted E.; Kennett, James P.; Kennett, Douglas J.; Moore, Andrew M. T.; Hillman, Gordon C.; Tankersley, Kenneth B.; Goodyear, Albert C. (4 de junho de 2013). «Evidence for deposition of 10 million tonnes of impact spherules across four continents 12,800 y ago». Proceedings of the National Academy of Sciences of the United States of America. 110 (23): E2088–2097. ISSN 1091-6490. PMID 23690611. doi:10.1073/pnas.1301760110
158. Wood TC, Cavanaugh DP (2001). A baraminological analysis of subtribe Flaveriinae (Asteraceae: Helenieae) and the origin of biological complexity. Origins. 52:7-27.
159. Wood TC, Wise KP, Sanders R, Doran N (2003). A Refined Baramin Concept. Occas. Papers of the BSG. 3:1-14.
160. Wood TC (2001). BDIST software, v. 1.0. Center for Origins Research and Education, Bryan College. Distributed by the author.
161. Wood TC (2002). A baraminology tutorial with examples from the grasses (Poaceae). Journal of Creation. 16(1):15–25.
162. Wood TC (2005). Visualizing baraminic distances using classical multidimensional scaling. Origins. 57:9-29.
163. Wood TC (2010). Baraminological Analysis Places Homo habilis, Homo rudolfensis, and Australopithecus sediba in the Human Holobaramin. Answers Research Journal 3:71–90.
164. Wood TC (2011). Using creation science to demonstrate evolution? Senter’s strategy revisited. Journal of Evolutionary Biology. 24(4):914–918.
165. Woodmorappe J (2000). The Karoo vertebrate nonproblem: 800 billion fossils or not. CEN Technical Journal. 14(2):47-49.
166. Woodruff DS (1980). Evolution: The Paleobiological View. Science. 208(4445):716-717.
167. Zhang, Y. P.; Yang, J. W.; Liu, Yi; Fan, T. S.; Luo, X. B.; Yuan, G. L.; Zhang, P. F.; Xie, X. F.; Song, X. Y. (June 2016). «Development of the radial neutron camera system for the HL-2A tokamak». The Review of Scientific Instruments. 87 (6). 063503 páginas. ISSN 1089-7623. PMID 27370450. doi:10.1063/1.4953109
168. Zellner, Nicolle E. B. (1 de setembro de 2017). «Cataclysm No More: New Views on the Timing and Delivery of Lunar Impactors». Origins of Life and Evolution of Biospheres (em inglês). 47 (3): 261–280. ISSN 0169-6149. doi:10.1007/s11084-017-9536-3
169. Zimmerman EC (1960). Possible evidence of rapid evolution in Hawaiian moths. Evolution. 14:137-138.
Martini, I. Peter; Baker, Victor R.; Garzón, Guillermina (5 de março de 2009). Flood and Megaflood Processes and Deposits: Recent and Ancient Examples (Special Publication 32 of the IAS) (em inglês). [S.l.]: John Wiley & Sons. ISBN 9781444304305 Disponível em https://books.google.com.br/books?hl=pt-BR&lr=&id=57wX5svvh8AC&oi=fnd&pg=PA3&ots=yPHV0d7eME&sig=cF1sJVvf1prTkt05DpnKRT2tqIw#v=onepage&q&f=false acessado em 16/12/2017
Sodré, GBN.; Souza Junior.; 2010 «Tempo de Formação: do Cambriano ao Pleistoceno | pt-brAdam's Apple». www.adamsapple.com.br. Consultado em 29 de novembro de 2017
Saitta, Evan T.; Rogers, Christopher S.; Brooker, Richard A.; Vinther, Jakob (1 de outubro de 2017). «Experimental taphonomy of keratin: a structural analysis of early taphonomic changes». PALAIOS: 647–657. ISSN 0883-1351. doi:10.2110/palo.2017.051
Axe, Douglas D.; Dixon, Brendan W.; Lu, Philip (4 de junho de 2008). «Stylus: a system for evolutionary experimentation based on a protein/proteome model with non-arbitrary functional constraints». PloS One. 3 (6): e2246. ISSN 1932-6203. PMID 18523658. doi:10.1371/journal.pone.0002246«Signature in the Cell « ASA Book Discussion». www.asa3online.org (em inglês). Consultado em 13 de outubro de 2017
Weber BH (1998) Emergence of life and biological selection from the perspective of complex systems dynamics. In: Van De Vijver G, Sathe SN, Delpos M (eds) Evolutionary Systems: Biological and Epistemological Perspectives on Selection and Self-Organization, pp 59–66. Kluwer, Dordrecht.
Kauffman SA (1993) The Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press, New York.
Morowitz H (1992) Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis. Yale University Press, New Haven.
Weber BH (2010) What is Life? Defining life in the context of emergent complexity. Origins of Life and Evolution of Biospheres 40: 221–229.
Schmitz, B. , Tassinari, M. & Peucker-Ehrenbrink, B. Uma chuva de condritas comuns no Ordoviciano adiantado . Planeta Terra. Sci. Lett. 194 , 1 - 15 ( 2001 ).
Schmitz, B., Tassinari, M. & Peucker-Ehrenbrink, B. A rain of ordinary chondrites in the early Ordovician. Earth Planet. Sci. Lett. 194, 1–15 (2001).
NATURE ASTRONOMY 1, 0035 (2017) | DOI: 10.1038/s41550-016-0035 | www.nature.com/natureastronomy 1
LETTERS
PUBLISHED: 23 JANUARY 2017 | VOLUME: 1 | ARTICLE NUMBER: 0035
© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.
Rare meteorites common in the Ordovician period
Rare meteorites common in the Ordovician period (PDF Download Available). Available from: https://www.researchgate.net/publication/312647580_Rare_meteorites_common_in_the_Ordovician_period [accessed Nov 21 2017]. «Rare meteorites common in the Ordovician period (PDF Download Available)». ResearchGate (em inglês). Consultado em 21 de novembro de 2017







