
ON THE ORIGIN OF STASIS
BY MEANS OF NATURAL PROCESSES
By
Art Battson
http://www.veritas-ucsb.org/library/battson/stasis/home.html
Tradução, inserções e comentários de Sodré GB Neto entre [ ]
“Estou bem ciente de que dificilmente há um único ponto discutido neste volume em que os fatos não possam ser aduzidos, muitas vezes aparentemente levando a conclusões diretamente opostas àquelas a que cheguei. Um resultado justo poderia ser obtido apenas declarando e equilibrando totalmente os fatos em ambos os lados de cada questão, e isso não pode ser feito aqui. ” Charles Darwin em "A origem das Espécies",1859
Charles Darwin estava bem ciente de que os cientistas podiam chegar a conclusões diretamente opostas às apresentadas em sua "Origem das Espécies" . Embora sua teoria pudesse explicar a pequena mudança evolutiva e a diversidade de tentilhões, Darwin sabia que tinha que ignorar virtualmente a história natural da vida na terra para manter qualquer esperança de explicar a origem dos filos e a maior disparidade entre artrópodes e antropólogos.
A teoria darwiniana está em conflito com as características mais proeminentes da história natural da Terra. Em primeiro lugar, a geologia não fornece as formas de transição que a teoria de Darwin exige. Em 1859, o conflito com a paleontologia era a objeção mais séria à teoria e, com o passar dos anos, a lacuna entre os dados e a teoria só aumentou; hoje os cientistas reconhecem menos formas de transição do que Darwin pensava que existiam. As espécies normalmente surgem de repente e "totalmente formadas".
O segundo conflito entre o darwinismo e a história natural é o fenômeno da estase. A geologia revela a estabilidade das formas, em vez de sua transformação gradual em planos corporais substancialmente diferentes. A estabilidade dos táxons superiores em particular sugere a existência de processos naturais que evitam que grandes mudanças evolutivas ocorram em uma base gradual passo a passo .
Finalmente, o padrão difuso de sucessão geológica é sistematicamente retrógrado em relação ao previsto pela teoria. A teoria darwiniana prevê que o acúmulo gradual de mudanças evolutivas menores e a diversidade crescente dos táxons inferiores devem, em última análise, produzir as profundas diferenças entre os planos corporais principais e a disparidade dos táxons superiores[ que era apenas conceitos abstratos organizacionais dos pais da taxonomia e de uma hora pra outra virou graus mais distante ou mais próximos de parentesco de uma evolução sem limites] . A diversidade deve preceder a disparidade. A sucessão geológica revela o oposto: a disparidade precede a diversidade. Os principais temas ou planos corporais aparecem repentinamente na história da vida apenas para serem seguidos por variações sobre esses temas pré-existentes . A história natural da vida na Terra é sistematicamente de cima para baixo, não de baixo para cima como a teoria darwiniana prevê.
Se Darwin tivesse desenvolvido uma teoria para explicar os dados empíricos da história natural, ele deveria ter chegado a conclusões diretamente opostas. Ele deveria ter desenvolvido uma teoria para explicar por que as espécies não mudam 1 gradualmente e se transformam em planos corporais substancialmente diferentes em uma base gradual passo a passo .
O fenômeno de estase e a estabilidade dos planos corporais principais é baseado em uma abundância de dados e nossas teorias que descrevem o mundo natural devem explicar esses dados. A evidência empírica sugere a necessidade de desenvolver uma teoria que seja baseada na história natural, em vez de uma que deva explicar suas características principais. Embora a teoria neodarwiniana ajude a explicar mudanças evolutivas menores, uma teoria de “macrostase” precisa ser desenvolvida para explicar a estabilidade dos planos corporais principais.
Devemos primeiro compreender as regras comuns de estabilidade e os padrões difusos da história natural antes de podermos especular sobre a origem dos principais planos corporais. Devemos também compreender que, em última análise, as questões das origens são metafísicas. As questões de microevolução e macrostase, entretanto, são claramente empíricas.
Conflitos entre Darwin e Paleontologia
Darwin viu a evolução como um processo lento e imponente. Ele imaginou organismos se transformando gradualmente de uma espécie para outra ao longo de imensos períodos de tempo. A evolução, ele acreditava, tinha que ocorrer por meio de “infinitamente numerosos elos de transição” formando “os melhores passos graduados.” Darwin era um adepto estrito do gradualismo e da noção de que “a natureza não dá saltos”. Ele explicou isso muito claramente em sua Origem das Espécies :
'Se pudesse ser demonstrado que existe qualquer órgão complexo que não pudesse ter sido formado por numerosas e sucessivas modificações leves, minha teoria seria totalmente destruída (Darwin, 1859, p. 219)".
Havia um grande obstáculo para essa visão da vida: a evidência fóssil. Em um capítulo intitulado "Sobre a imperfeição do registro geológico", ele admite prontamente:
... O número de variedades intermediárias, que existiram anteriormente na terra, (deve) ser verdadeiramente enorme. Por que então nem toda formação geológica e todo estrato estão cheios desses elos intermediários? A geologia certamente não revela nenhuma cadeia orgânica finamente graduada; e esta, talvez, seja a objeção mais óbvia e mais grave que pode ser levantada contra minha teoria (Darwin, 1859, p. 292).
Apesar dos sérios problemas apresentados pelas evidências geológicas, Darwin acreditava que a passagem do tempo revelaria o enorme número de formas de transição que sua teoria exigia. No entanto, não foi esse o caso. David Raup, ex-curador de geologia do Field Museum of Natural History de Chicago, colocou desta forma:
Bem, estamos agora cerca de 120 anos depois de Darwin, e o conhecimento do registro fóssil foi muito expandido ... ironicamente, temos ainda menos exemplos de transição evolutiva do que na época de Darwin. Com isso, quero dizer que alguns dos casos clássicos de mudança darwiniana no registro fóssil, como a evolução do cavalo na América do Norte, tiveram que ser descartados ou modificados como resultado de informações mais detalhadas ... (Raup, 1979 )
O caso paleontológico contra o gradualismo era sério na época de Darwin e só piorou as coisas. Stephen Jay Gould, professor de geologia e paleontologia da Universidade de Harvard, explica:
A história da maioria das espécies fósseis inclui duas características particularmente inconsistentes com o gradualismo:
1) Estase - a maioria das espécies não exibe nenhuma mudança direcional durante sua permanência na Terra. Eles aparecem no registro fóssil com a mesma aparência de quando desaparecem; a mudança morfológica é geralmente limitada e sem direção;
2) Aparecimento repentino - em qualquer área local, uma espécie não surge gradualmente pela transformação contínua de seus ancestrais; ele aparece de uma só vez e "totalmente formado" (Gould, 1977)
Gould honestamente admite que a síntese neodarwiniana não é suportada pela evidência fóssil e “está efetivamente morta, apesar de sua persistência como ortodoxia de livro didático” (Gould, 1980).
Embora a estase seja a característica dominante da história da vida, exceções ao padrão geral de estase podem ser citadas. Exemplos de séries de transição podem ser encontrados em níveis taxonômicos mais baixos. Em níveis taxonômicos mais altos, no entanto, as sequências de transição variam de escassas a inexistentes. A evidência de gradualismo entre filos, classes e até mesmo ordens é inexistente ou é muito contestada. Certamente, nenhum padrão difundido de gradualismo existe. George Gaylord Simpson [muito elogiado por Gould devido seu imenso trabalho] reconheceu isso décadas atrás, ao descrever a situação nestes termos:
"Isso é verdade para todas as trinta e duas ordens de mamíferos ... Os primeiros e mais primitivos membros conhecidos de cada ordem já possuem os caracteres ordinais básicos e, em nenhum caso, é conhecida uma sequência aproximadamente contínua de uma ordem para outra. Na maioria dos casos, a quebra é tão acentuada e a lacuna tão grande que a origem do pedido é especulativa e muito disputada ...
Essa ausência regular de formas de transição não se limita aos mamíferos, mas é um fenômeno quase universal, como há muito tempo foi observado pelos paleontólogos. É verdade para quase todas as classes de animais, tanto vertebrados quanto invertebrados ... é verdade para as classes e para os principais filos animais e, aparentemente, também é verdade para categorias análogas de plantas" (Simpson, 1944).
Pesquisas recentes sobre a origem dos táxons superiores confirmam o que os paleontólogos já sabem há décadas.
Táxons reconhecidos como ordens durante a transição (pré-cambriano-cambriano) aparecem principalmente sem conexão com um clado ancestral por meio de um fóssil intermediário. Esta situação também é verdadeira para a maioria das ordens de invertebrados durante o Fanerozóico remanescente. Não há cadeias de taxa que conduzam gradualmente de uma condição ancestral ao novo tipo de corpo ordinal. As ordens, portanto, aparecem como subdivisões bastante distintas de classes, em vez de segmentos em algum tipo de continuum morfológico (Valentine, Awramik, Signor e Sadler, 1991).
A origem das classes e filos constitui uma dificuldade ainda maior para a teoria neodarwiniana . Para agravar o problema, está a pequena janela de tempo disponível para a origem da grande maioria dos filos. Pesquisas recentes reduziram o “Big Bang da biologia” a alguns milhões de anos. (Ver Kerr, 1993 e Bowring et al, 1993.) Quando se compara o período de tempo que os processos darwinianos levaram para modificar o bico de um tentilhão com o período de tempo, virtualmente todos os principais planos corporais apareceram na explosão cambriana, torna-se difícil acreditar que um acúmulo gradual de mudanças microevolutivas teve muito a ver com a origem dos taxa superiores.
A evolução darwiniana prevê a presença regular de formas de transição. O registro fóssil revela sua ausência regular. Também revela um fenômeno natural que até recentemente era praticamente ignorado pelos paleontólogos. Esse fenômeno é a estase. A tragédia do darwinismo é que ele impediu o progresso da ciência ao desviar a atenção dos biólogos e paleontólogos dos dados empíricos e distraí-los do desenvolvimento de teorias que explicam o fenômeno natural difuso da estase. Por mais de 130 anos, cientistas que trabalham dentro do paradigma darwiniano têm tentado desenvolver teorias para explicar dados que, no nível macro, não existem (ver Figura 1).
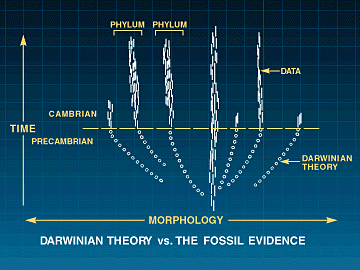
Figura 1: Teoria Darwiniana vs. o Registro Fóssil
A teoria darwiniana tenta explicar a ancestralidade comum de todas as espécies por meio da transformação gradual dos principais planos corporais. Essa teoria está em oposição à evidência fóssil e aos padrões difusos da história natural.
Estima-se que 75 a 100 filos aparecem de forma explosiva na base do Cambriano. Evidências fósseis sugerindo sua ancestralidade comum não são encontradas em rochas pré-cambrianas. Uma Teoria Geral de Macrostasia é necessária para explicar os dados fósseis e a estabilidade dos táxons superiores.
Escrevendo na introdução à reedição de Origem das Espécies , em 1956 , WR Thompson comentou:
O sucesso do darwinismo foi acompanhado por um declínio na integridade científica. Isso já é evidente nas declarações imprudentes de Haeckel e na argumentação astuta, tortuosa e histriônica de TH Huxley ...
Para estabelecer a continuidade exigida pela teoria, argumentos históricos são invocados, embora faltem evidências históricas. Assim, são engendradas aquelas frágeis torres de hipóteses baseadas em hipóteses, onde fato e ficção se confundem em uma confusão inextricável (Thompson, 1956).
Os dados fósseis mostram claramente padrões de estase, em vez de sequências evolutivas principais, e é para esse fenômeno que os cientistas devem voltar sua atenção. Como Niles Eldredge e Stephen Jay Gould colocaram: “Stasis são dados” (Gould, 1991). Os cientistas não podem perder de vista essas abundantes evidências históricas. Gould descreveu recentemente a importância de compreender a estase nestes termos:
... devemos entender que nada acontece na maioria das vezes - e não acontece porque nossas histórias não admitem esse tema - se esperamos compreender a dinâmica da mudança evolutiva. (Esta frase pode parecer contraditória, mas não é. Para saber as razões para mudanças infrequentes, é preciso entender as regras comuns de estabilidade.) O Burgess Shale nos ensina que, para a história dos designs anatômicos básicos, quase tudo aconteceu em o momento geológico imediatamente anterior, e quase nada em mais de 500 milhões de anos desde (Gould, 1988).
É inteiramente concebível que os processos naturais por si só sejam insuficientes para superar o que Gould chamou de “as regras comuns de estabilidade”. Kurt Wise, um ex-aluno de doutorado de Gould, sugeriu que pode haver pelo menos quatro níveis distintos de estase: nível molecular, nível de população, nível de espécie e estase de nível de táxon superior . Embora Wise acredite que os três primeiros níveis de estase são violáveis[devido a hipervariabilidade, característica que as espécies mães ancestrais possuem e isso pode ser verificado até hoje], ele aponta que pode haver um mecanismo que impede a mudança em taxa superiores que é inviolável. Rejeitando as suposições metafísicas de Gould, Wise conclui que provavelmente existem processos naturais que evitam que grandes mudanças evolutivas transformem o baramin, ou tipos originalmente criados, em planos corporais significativamente diferentes:
Provavelmente é apenas a estase no nível de taxa mais elevada que é válida e difere qualitativamente dos outros níveis de estase. Apenas táxons mais altos carecem de evidências demonstráveis de mudança ... Estagnação no nível do táxon mais alto poderia ser o resultado do que poderia ser chamado de Estase Baramínica - a restrição permanente dos organismos sob condições naturais para permanecer dentro dos limites de seus baramins (Wise, 1991) .
O conceito de baramin é sinônimo do conceito de “espécie criada” e é um anátema para os cientistas que acreditam que a origem e a diversidade da vida devem ser atribuídas a processos puramente mecanicistas. O objetivo da ciência, no entanto, não deve ser desenvolver um “relato da criação” naturalista em uma tentativa de explicar a origem e a diversidade de toda a vida por meios puramente materialistas. Em vez disso, o objetivo da ciência deve ser descrever com mais precisão os padrões e fenômenos difundidos encontrados na natureza, mesmo que esses processos naturais impeçam a ocorrência de grandes mudanças evolutivas. A ciência precisa de uma diversidade de idéias livres do naturalismo filosófico. Os cientistas precisam de uma teoria para explicar o fenômeno do nível de táxon mais alto estase e uma teoria para explicar por que as espécies não parecem evoluir gradualmente para algo substancialmente diferente.
Conflitos entre Darwin e sucessão geológica
Antes de A Origem das Espécies foi escrita, a escala de tempo geológica em sua forma moderna já estava totalmente desenvolvida. Na verdade, a escala de tempo baseada em fósseis foi construída por geólogos criacionistas (Raup, 1981). Os cientistas da época de Darwin não igualavam a sucessão geológica à evolução, nem deveríamos hoje. Um dos motivos é que a sucessão geológica não revela como as novas espécies passaram a existir, apenas quando . Outra razão é que a ordem de aparência encontrada no registro fóssil é, como regra geral, sistematicamente retroativa em relação às principais previsões da teoria darwiniana.
A sucessão geológica é freqüentemente considerada como evidência primária do fato da evolução. As amebas aparecem antes dos peixes e os peixes antes dos anfíbios. Na visão popular, essa sucessão do simples ao complexo é uma evidência da evolução. A sucessão geológica, entretanto, não procede necessariamente do simples ao complexo. Por exemplo, os trilobitas, entre os artrópodes mais avançados, são os primeiros artrópodes a aparecer no registro geológico. Darwin lamentou a complexidade do olho dos vertebrados, confessando que:
“supor que o olho ... poderia ter sido formado por seleção natural, parece, eu confesso livremente, absurdo no mais alto grau possível” (Darwin, 1859, p. 217 )
No entanto, alguns cientistas acreditam que os olhos esquizocroais de alguns trilobitas possuíam os sistemas ópticos mais sofisticados já utilizados por qualquer organismo(Levi-Setti, 1975). David Raup afirmou que os trilobitas “... usaram um design óptico que exigiria um engenheiro óptico bem treinado e imaginativo para se desenvolver hoje ... (Raup, 1979)”. Os primeiros metazoários eram tudo menos simples. A complexidade dos primeiros metazoários, entretanto, é apenas uma ruga na ironia do darwinismo. A verdadeira ironia está na ordem sistemática geral de aparecimento dos táxons no registro fóssil.
A vida é complexa demais para que nossas teorias prevejam os planos corporais específicos que deveriam surgir como resultado da evolução. Tanto o gradualismo darwiniano quanto o equilíbrio pontuado, entretanto, predizem que um padrão de ramificação sistemático deveria ser evidente se toda a vida surgiu de um ancestral comum (ver Figuras 2 e 3).
Ambos os modelos são baseados em espécies intra ou interespécies relativamente de pequenas mudanças e, portanto, prevêem que numerosas espécies transicionais devem vir à existência, as quais gradualmente divergem para produzir a disparidade dos taxa superiores. [Lembrando que podemos ver estabilidade e sucesso reprodutivo na biodiversidade de hoje também, ou seja, não há desculpa de que transicionais sejam pouco duráveis no tempo para que não sejam encontradas no registro fóssil]
Ambos os modelos prevêem que, à medida que novas espécies surgem e a distância morfológica entre elas aumenta, novos gêneros e, eventualmente, novas famílias devem aparecer. Conforme ocorre o aumento da diversidade, novas ordens, novas classes e, finalmente, novos filos devem surgir. Em suma, a diversidade deve preceder a disparidade. O aumento gradual da diversidade dos táxons inferiores deve resultar, em última instância, na disparidade dos táxons superiores e no aparecimento de novos planos corporais importantes. O padrão de sucessão geológica predito por ambas as teorias evolutivas é de baixo para cima: de espécies a gêneros, a famílias, a ordens, a classes, a filos.[Ou seja, a realidade é invertida pelas duas teorias gradualistas e pontualistas]
O padrão generalizado de sucessão geológica é contrário a essas teorias (ver Figura 4). A disparidade precede a diversidade. O aparecimento inicial de virtualmente todos os filos ocorre com diversidade de espécies muito baixa. A origem dos planos corporais principais não é o resultado da diversidade crescente dos táxons inferiores; o padrão geral não é de baixo para cima. Em vez disso, o padrão dominante é de cima para baixo, ao contrário da teoria. Como os paleontólogos Douglas Erwin, James Valentine e John Sepkoski descrevem a situação:
O registro fóssil sugere que o pulso principal de diversificação dos filos ocorre antes das classes, as classes antes das ordens e as ordens antes das famílias. Isso não quer dizer que cada táxon superior se originou antes das espécies (cada filo, classe ou ordem continha pelo menos uma espécie, gênero, família, etc. após o aparecimento), mas os táxons superiores não parecem ter divergido por meio de um acúmulo de taxa inferior (Erwin, Valentine e Sepkoski, 1988).

Figura 2: Gradualismo filético
Através da transformação continuamente gradual das espécies, este modelo prevê que a diversidade crescente dos taxa inferiores deve preceder a disparidade dos taxa superiores.
Figura 3: Equilíbrio pontuado
Sob este modelo, a mudança evolutiva está confinada a intervalos de tempo mais curtos e pequenas populações isoladas para dar conta da falta de formas de transição entre os táxons inferiores. Pontuações no nível do táxon inferior , entretanto, requerem numerosas etapas de transição para produzir a disparidade dos táxons superiores. Este modelo também prevê que o aumento da diversidade dos táxons mais baixos deve preceder a disparidade dos táxons mais altos.

Figura 4: a evidência fóssil
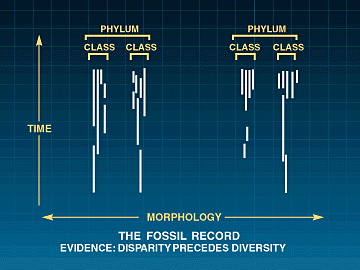
Ao contrário do gradualismo darwiniano e da teoria do equilíbrio pontuado, a grande maioria dos filos aparecem abruptamente com baixa diversidade de espécies. A disparidade dos táxons mais altos precede a diversidade dos táxons mais baixos.
Em outro artigo, Valentine e Erwin revisam as hipóteses quanto ao modo de origem dos planos do corpo animal quanto à consistência com a evidência fóssil. Eles concluem que tanto o gradualismo darwiniano quanto o equilíbrio pontuado são inadequados para explicar o aparecimento de planos corporais de invertebrados e suas principais modificações:
Os modelos que consideramos são de três tipos: aqueles que extrapolam os processos de especiação para contabilizar táxons superiores via divergência, aqueles que invocam a seleção entre as espécies e aqueles que enfatizam que muitos táxons superiores se originaram como novas linhagens por direito próprio, não apenas como uma conseqüência de processos em nível de espécie . É nessa última classe de modelo que acreditamos que o registro favorece (Valentine e Erwin, 1985, p. 71).
Se grandes populações evoluíram gradualmente, deveria haver evidência inconfundível no registro fóssil, mas simplesmente não foi encontrado.
... muitas das grandes populações deveriam ter sido preservadas, mas nós simplesmente não as encontramos. Portanto, são necessárias pequenas populações, mas aqui também existem dificuldades. As populações devem permanecer pequenas (e não detectadas) e evoluir de forma constante e consistente em direção ao plano corporal que constitui a base de um novo filo (ou classe). Isso é pedir muito. Mutações deletérias tendem a se acumular em pequenas populações para formar cargas genéticas que a seleção pode não ser capaz de controlar. Os modos adaptativos intermediários estáveis não podem ser invocados como um recurso regular, uma vez que nos deparamos novamente com o problema de onde estão seus restos. Podemos imaginar vastas matrizes dessas pequenas populações se espalhando contínua e incessantemente no espaço adaptativo. Vastas matrizes deveriam ter produzido pelo menos alguns fósseis também permanecem. Talvez uma dificuldade ainda maior seja a exigência de que esses arranjos de linhagens mudem ao longo de um curso bastante reto e verdadeiro - viagens morfológicas laterais ou desvios de qualquer freqüência deveriam prolongar o tempo de origem dos táxons superiores além do que parece estar disponível. Por que deveria um processo oportunista e improvisado estabelecer tal curso e mantê-lo por tanto tempo com sucesso entre tantas linhagens?
Concluímos que a extrapolação de taxas microevolucionárias para explicar a origem de novos planos corporais é possível, mas não está de acordo com a evidência primária (Valentine e Erwin, 1985, pp. 95, 96).
O modelo de equilíbrio pontuado ou seleção de espécies tenta explicar a falta de evidências, baseando-se principalmente na evolução de pequenas populações isoladas que teriam uma chance reduzida de deixar um registro fóssil. Este cenário tem suas dificuldades, no entanto, como Valentine e Erwin apontam:
A rapidez necessária para a mudança implica alguns passos grandes ou muitos passos menores e excessivamente rápidos. Passos grandes equivalem a saltos e levantam os problemas de barreiras de adequação; os pequenos passos devem ser numerosos e envolver os problemas discutidos na microevolução. Os períodos de estase levantam a possibilidade de que a linhagem entraria no registro fóssil, e reiteramos que não podemos identificar nenhuma das formas intermediárias postuladas. Finalmente, o grande número de espécies que devem ser geradas para formar um pool a partir do qual a linhagem bem-sucedida é selecionada não estão em lugar algum. Concluímos que a probabilidade de que a seleção de espécies seja uma solução geral para a origem de táxons superiores não é grande, e que nenhuma das teorias contrárias de mudança evolutiva no nível de espécie,
Essa evidência confirma ainda mais a proposição de que mudanças evolutivas menores de nível inferior não podem ser extrapoladas para explicar as mudanças evolutivas maiores. Isso parece ser verdade tanto para a tartaruga quanto para a lebre, o gradualismo darwiniano e o equilíbrio pontuado. A tartaruga é lenta demais para explicar a evidência fóssil e a lebre passa muito tempo em estase.
Darwin admitiu que a evidência geológica era a "objeção mais óbvia e mais grave que pode ser levantada contra minha teoria". Na época, ele estava principalmente preocupado com a falta de formas de transição no registro fóssil. Hoje, essas preocupações são agravadas por menos formas de transição que Darwin tinha em sua época, e pela ordem de sucessão geológica sistematicamente invertida , Darwin foi ironicamente profético ao afirmar que os fatos podem levar a conclusões diretamente opostas àquelas a que chegou.
Roger Lewin descreve ainda a origem da maioria dos planos corporais importantes no relatório da Science Research News, "A Lopsided Look at Evolution":
Descrita recentemente como "o evento evolucionário mais importante durante toda a história dos Metazoa", a explosão cambriana estabeleceu virtualmente todas as principais formas do corpo animal - Bauplane ou filos - que existiriam depois disso, incluindo muitos que foram 'eliminados' e se extinguiram . Comparado com os cerca de 30 filos existentes, algumas pessoas estimam que a explosão do Cambriano pode ter gerado até 100. A inovação evolutiva da fronteira pré-cambriana / cambriana foi claramente extremamente amplo: “sem precedentes e insuperável”, como James Valentine, da Universidade da Califórnia, Santa Bárbara, recentemente colocou (Lewin, 1988).
Lewin então fez a importante pergunta:
"Por que, em períodos subsequentes de grande atividade evolutiva, quando incontáveis espécies, gêneros e famílias surgiram, não houve novos planos do corpo animal produzidos, nem novos filos?" (Lewin, 1988).
Se a teoria neodarwiniana é verdadeira, por que deveria o Cambriano conter um número maior de plantas corporais do que existe hoje, particularmente com tão baixa diversidade de espécies? As Figuras 5 e 6 ilustram graficamente a situação.
A pergunta de Lewin nos leva a uma pergunta ainda mais importante. Que mecanismos evitaram que grandes mudanças evolutivas ocorressem nos últimos 500 milhões de anos? Por que a origem dos filos parece ter parado primeiro, seguido pelas classes e depois pelas ordens?
Uma explicação bastante convincente que foi oferecida para o padrão é baseada no fato de que qualquer grande novidade que surja com relativa rapidez por meio de processos não seletivos provavelmente será mal adaptada. Quanto mais inovador for o plano do corpo, mais suscetível ele será à eliminação pela competição e, portanto, mais espaço adaptativo será necessário para que ele se estabeleça. Em suma, a competição tende a inibir o estabelecimento de táxons superiores. A seleção natural tende a inibir grandes mudanças evolutivas.

Figura 5: A Explosão Cambriana
O aparecimento súbito de cerca de 100 planos corporais distintos com diversidade extremamente baixos espécies suporta a conclusão de que nem a evolução darwiniana gradual nem menores de nível taxon pontuações pode explicar a origem da taxa mais elevada e as grandes planos corporais. Na história da vida na Terra, a disparidade geralmente precede a diversidade.
Figura 6: o presente

“Podemos reconhecer um fato central e surpreendente da história da vida - diminuição acentuada na disparidade seguida por um aumento notável na diversidade dentro dos poucos designs sobreviventes.” (Stephen Jay Gould, 1989)
Sobre a origem da estase por meio da seleção natural
Charles Darwin tornou plausível o conceito de grande mudança evolutiva ao convencer os cientistas de que a seleção natural poderia ser responsável pelo surgimento do design na natureza (Horan, 1979). Ele nunca teria considerado a evolução um fato sem uma teoria plausível de como ela poderia ocorrer. O próprio título de seu livro reflete a importância de um mecanismo evolucionário. Embora muitas evidências tenham sido citadas a favor da macroevolução, como era antes de 1859, tais evidências, na opinião do próprio Darwin, seriam insatisfatórias sem um mecanismo:
Ao considerar a Origem das Espécies, é perfeitamente concebível que um naturalista, refletindo sobre as afinidades mútuas dos seres orgânicos, sobre suas relações embriológicas, sua distribuição geográfica, sucessão geológica e outros fatos, possa chegar à conclusão de que cada espécie teve não foi criado independentemente, mas descendeu, como variedades, de outras espécies. No entanto, tal conclusão, mesmo que bem fundamentada, seria insatisfatória, até que se mostrasse como se modificaram as inúmeras espécies que habitam este mundo, de forma a adquirir aquela perfeição de estrutura e coadaptação que justamente nos excita a admiração ( Darwin, 1859, p. 66).
Ao considerar a origem dos filos e a estabilidade dos principais planos corporais, é perfeitamente concebível que os cientistas, refletindo sobre as evidências empíricas da história da vida na Terra, possam chegar à conclusão de que existem mecanismos que impedem que grandes mudanças evolutivas ocorram em uma base gradual passo a passo . A estase é um fato da vida e devem existir mecanismos que explicam isso. Ironicamente, a própria seleção natural pode desempenhar um papel importante no fenômeno da estase no nível do táxon superior .
Darwin teve seus críticos. Em seu livro Darwinism: The Refutation of a Myth , Soren Lovtrup aponta que “alguns críticos se voltaram contra os ensinamentos de Darwin por razões religiosas, mas eles eram uma minoria; a maioria de seus oponentes ... argumentou em uma base completamente científica. ” Ele continua explicando:
... as razões para rejeitar a proposta de Darwin foram muitas, mas antes de tudo que muitas inovações não podem vir a existir através da acumulação de muitos pequenos passos e, mesmo que possam, a seleção natural não pode realizá-lo, porque os estágios incipientes e intermediários não são vantajoso (Lovtrup, 1987).
Talvez o mais formidável dos críticos de Darwin tenha sido St. George Mivart. Seu principal livro, On the Genesis of Species , visava a noção de que a seleção natural poderia ser responsável pelo acúmulo de estágios incipientes de estruturas úteis (Mivart, 1871). Stephen Jay Gould observa que Darwin ofereceu fortes, embora relutantes, elogios e levou Mivart muito mais a sério do que qualquer outro crítico ... Mivart reuniu, e ilustrou "com admirável arte e força" (palavras de Darwin), todas as objeções à teoria da seleção natural - "um formidável array ”(palavras de Darwin novamente). No entanto, um tema particular, incentivado com atenção especial por Mivart, se destacou como a peça central de sua crítica. Continua a ser hoje a principal pedra de tropeço entre os escrutinadores atenciosos e amigáveis do darwinismo. Nenhuma outra crítica parece tão perturbadora, tão óbvia e evidentemente “certa” (contra uma afirmação darwiniana que parece intuitivamente paradoxal e improvável).
Mivart concedeu a esta crítica um capítulo separado em seu livro, logo após a introdução. Ele também deu um nome, lembrado desde então. Ele chamou de "A incompetência de
frequência da "Seleção Natural" para explicar os estágios iniciais de estruturas úteis:
" Se esta frase soa como um bocado, considere a tradução fácil: podemos compreender prontamente como estruturas complexas e totalmente desenvolvidas funcionam e devem a sua manutenção e preservação à seleção natural - uma asa, um olho, a semelhança de um bitterno com um galho ou de um inseto para um pedaço de pau ou folha morta. Mas como você vai do nada para algo tão elaborado se a evolução deve prosseguir através de uma longa sequência de estágios intermediários, cada um favorecido por seleção natural? Mas como você vai do nada para algo tão elaborado se a evolução deve prosseguir através de uma longa sequência de estágios intermediários, cada um favorecido por seleção natural? Você não pode voar com 2% de uma asa ou ganhar muita proteção. Como, em outras palavras, pode a seleção natural explicar esses estágios incipientes de estruturas que só podem ser usados (como nós agora observamos) de uma forma muito mais elaborada (Gould, 1985)?

Gould prossegue apontando que, entre as dificuldades da teoria darwiniana, “um ponto se destaca muito acima do resto: o dilema dos estágios incipientes. Mivart identificou esse problema como o principal e continua sendo até hoje. ”
Existem inúmeros exemplos de organismos com sistemas de componentes altamente especializados e interdependentes, todos os quais devem ser integrados antes de serem funcionais e oferecerem qualquer vantagem seletiva. A natureza abunda em tais sistemas de "complexidade irredutível" . Embora possa haver alguns exemplos onde certos componentes assumem alguma função pré-adaptativa, esses casos podem ser considerados como exceções a uma regra mais geral. Via de regra, qualquer subconjunto dos componentes seria um fardo para um organismo e, portanto, seria eliminado pela seleção natural. Tome por exemplo o mecanismo sensorial e motor da bactéria comum, Escherichia coli , um organismo procariótico unicelular relativamente simples.
[Existem diversos motores que podemos chamar de "modificadores" e/ou sub-especiativos , atuando nas populações como por exemplo, deriva genética, fluxo gênico, epigenética, sistema imunológico, energias que atuam na hora do crossing over, e o motor das mutações sendo a maioria deletérias (entropia) que parece reger a todos os outros, onde tudo que for conservativo, atua dentro de sua plataforma de degeneração, entrópica, desorganizando cada vez mais informações genéticas, como se tivesse mais engrenagens que agissem em todos os outros motores. Assim descreve Sodré GB Neto em artigo científico ligado a diabetes:
"Toda ciência agronômica, botânica, zootecnista, médica veterinária e medicina humana já usa NGS para administrar as mutações que vão se acumulando nas populações[26] e em nossa vida como indivíduos, espermas[27], idade[28][29]pois a entropia ocorre em quaisquer sistemas, sobretudo complexos como é o sistema informacional [30] [31]genético[32] acumulando mutações cada vez mais[33]. Diante deste quadro cada vez mais ameaçador, onde "levando-se em conta a gravidade, em média, todas as pessoas do mundo tiveram uma redução de 11% em sua saúde geral entre 1990 e 2010 devido a doenças e lesões" [34], compreender a doença em sua raiz já deixou de ser um luxo acadêmico intelectual, mas uma urgente necessidade. Infelizmente percebemos que no Brasil a maioria dos diagnósticos de doenças ainda estão nos tempos das cavernas bem escuras, onde os profissionais apenas apalpam os problemas como cegos para a realidade das mutações genéticas (a maioria deletérias) cada vez mais presente:
"Além de herdar metade do genoma de cada um de nossos pais, nascemos com um pequeno número de novas mutações que ocorreram durante a gametogênese e pós-zigoticamente. Estudos recentes de sequenciamento de genoma e exoma de trios pais-filhos forneceram os primeiros insights sobre o número e a distribuição dessas mutações de novo na saúde e na doença, apontando para fatores de risco que aumentam seu número na descendência. Mutações de novo demonstraram ser a principal causa de distúrbios genéticos graves de início precoce, como deficiência intelectual, distúrbio do espectro do autismo e outras doenças do desenvolvimento. Na verdade, a ocorrência de novas mutações em cada geração explica por que esses distúrbios reprodutivamente letais continuam a ocorrer em nossa população. Estudos recentes também mostraram que as mutações de novo são predominantemente de origem paterna e que seu número aumenta com a idade paterna avançada.". [35]
Já está estabelecido ao contrário do que muitos acreditavam em mutações positivas e maioria neutras, que a maioria delas são deletérias[36]:
"A predominância de mutações deletérias sobre as benéficas está bem estabelecida. James Crow em ( 1997 ) afirmou: “Uma vez que a maioria das mutações, se elas têm algum efeito, são prejudiciais, o impacto geral do processo de mutação deve ser deletério”. Keightley e Lynch ( 2003 ) deram uma excelente visão geral dos experimentos de acumulação de mutações e concluíram que “... a grande maioria das mutações são deletérias. Este é um dos princípios mais bem estabelecidos da genética evolutiva, apoiado por dados genéticos moleculares e quantitativos. Isso fornece uma explicação para muitas propriedades genéticas importantes das populações naturais e de laboratório ”. Em ( 1995 ), Lande concluiu que de novas mutações são deletérias e, o resto é “quase neutro” (ver também Franklin e Frankham ( 1998 )). Gerrish e Lenski estimam a proporção de mutações deletérias para benéficas em um milhão para um (Gerrish e Lenski 1998b ), enquanto outras estimativas indicam que o número de mutações benéficas é muito baixo para ser medido estatisticamente (Ohta 1977 ; Kimura 1979 ; Elena et al . 1998 ; Gerrish e Lenski 1998a ). Estudos em diferentes espécies estimam que, além da seleção, a diminuição na aptidão de mutações é de 0,2-2 por geração, com declínio da aptidão humana estimado em (Ver Lynch 2016 ; Lynch et al. 1999 ). As estimativas sugerem que o recém-nascido humano médio tem aproximadamente 100 mutações de novo (Lynch 2016 ). A pesquisa usando modelos de população finita foi impulsionada pela necessidade de entender o impacto do acúmulo de mutações deletérias (chamadas de carga mutacional) em pequenas populações de espécies ameaçadas de extinção (ver Lande 1995 ; Franklin e Frankham 1998 ). De especial interesse é a carga mutacional na espécie humana, dada a seleção relaxada devido aos avanços sociais e médicos (Kondrashov 1995 ; Crow 1997 ; Lynch 2016 )".(Sodré GB Neto, 2020)
E a entropia uma vez percebida, nos aponta um passado cada vez com menor imperfeição das informações genéticas ].
Figura 7. Diagrama de blocos conceitual do mecanismo sensorial e motor da bactéria, E. coli .

O mecanismo sensorial e motor consiste em vários receptores que detectam inicialmente as concentrações de uma variedade de produtos químicos. Os componentes secundários extraem informações desses sensores que, por sua vez, são usados como entrada para um mecanismo de detecção de gradiente. A saída desse mecanismo é usada para acionar um conjunto de motores rotativos reversíveis alimentados por prótons de torque constante que transferem sua energia através de um trem de acionamento microscópico e impulsionam flagelos helicoidais (ver Figura 7). Este sistema altamente integrado permite que a bactéria migre a uma taxa de aproximadamente dez comprimentos corporais por segundo.
O Dr. Robert Macnab, da Universidade de Yale, concluiu uma importante revisão de 50 páginas desse mecanismo com as seguintes observações:
Como comentário final, podemos apenas maravilhar-nos com a complexidade em uma simples bactéria, do sistema motor e sensorial total que foi o assunto desta revisão e observar que nosso conceito de evolução por vantagem seletiva deve certamente ser uma simplificação excessiva. Que vantagem poderia derivar, por exemplo, de um “pré-lagelo” (significando um subconjunto de seus componentes) e, ainda assim, qual é a probabilidade de desenvolvimento “simultâneo” da organela em um nível em que se torna vantajoso? (Macnab, 1978)
Cada componente do flagelo sozinho (veja a Figura 8) é altamente especializado em sua função e é altamente improvável que muitos dos componentes, se algum, tivessem qualquer valor pré-adaptativo. Um subconjunto de componentes constituiria uma coleção de partes supérfluas que, de acordo com a teoria darwiniana, deveriam ser eliminadas pela seleção natural. Darwin apontou em The Descent of Man (Darwin, 1871) que a seleção natural agiria para preservar aqueles indivíduos que estivessem menos sobrecarregados com uma parte supérflua. Embora seja um dos organismos mais simples de nosso planeta, ele serve para ilustrar como a seleção natural contribui para o fenômeno da estase e ajuda a explicar o fato de que a maioria das espécies não surge gradualmente pela transformação constante de seus ancestrais.
Fatos, fósseis e filosofia
O clássico de Charles Darwin, Na Origem das Espécies por Meio da Seleção Natural , foi um argumento convincente e convincente para a ideia de que pequenas mudanças evolutivas poderiam ser extrapoladas para explicar a origem de todas as principais formas de vida por métodos puramente mecanicistas e materialistas processos. Escrevendo sobre a origem e o impacto do pensamento evolucionário, Douglas Futuyma colocou desta forma:
Darwin mostrou que as causas materiais são uma explicação suficiente não apenas para os fenômenos físicos, como Descartes e Newton mostraram, mas também para os fenômenos biológicos com todas as suas evidências aparentes de desígnio e propósito. Ao acoplar a variação não direcionada e sem propósito ao processo cego e indiferente da seleção natural, Darwin tornou supérfluas as explicações teológicas ou espirituais dos processos vitais. Junto com a teoria materialista da história e da sociedade de Marx e a atribuição de Freud do comportamento humano a influências sobre as quais temos pouco controle, a teoria da evolução de Darwin foi uma plataforma crucial na plataforma do mecanicismo e do materialismo - de grande parte da ciência, em resumo - que desde então, foi o palco da maior parte do pensamento ocidental (Futuyma, 1986).
As extrapolações de Darwin foram justificadas? A julgar pelas conclusões de muitos dos cientistas que participaram de uma das conferências mais importantes de biologia evolutiva nos últimos quarenta anos, a resposta provavelmente não é.
A questão central da conferência de Chicago foi se os mecanismos subjacentes à microevolução podem ser extrapolados para explicar os fenômenos da macroevolução. Correndo o risco de violentar as posições de algumas pessoas na reunião, a resposta pode ser dada como um claro Não (Lewin, 1980).
O artigo de Lewin sobre a conferência na revista AAAS, Science , descreveu alguns dos principais conflitos entre a moderna teoria neodarwiniana da evolução e as evidências empíricas:
A evolução, de acordo com a Síntese Moderna, se move em um ritmo majestoso, com pequenas mudanças se acumulando ao longo de períodos de muitos milhões de anos, gerando uma longa herança de linhagens em avanço constante, conforme revelado no registro fóssil. No entanto, o problema é que, de acordo com a maioria dos paleontólogos, a principal característica das espécies individuais no registro fóssil é a estase, não a mudança ...
Em uma admissão generosa, Francisco Ayala, uma figura importante na proposição da Síntese Moderna nos Estados Unidos, disse "Não teríamos previsto a estase da genética populacional, mas agora estou convencido do que dizem os paleontólogos de que pequenas mudanças não se acumulam" ( Lewin, 1980).
Se pequenas mudanças não se acumulam, se a estase é a principal característica das espécies individuais na história natural da vida, se a seleção natural inibe ou impede grandes mudanças evolutivas, se a ordem de aparecimento no registro geológico é sistematicamente retrógrado para as previsões darwinianas, e se os táxons superiores são tão descontínuos quanto parecem, por que os cientistas não desenvolvem teorias para explicar os limites naturais da mudança biológica? A resposta parece ser mais filosófica do que empírica. Embora a questão provavelmente levasse a uma descrição mais precisa da natureza, sem dúvida o prejudicaria .
A filosofia secular difusa de mecanicismo e materialismo que passou a dominar a ciência moderna. Embora a pergunta possa certamente levar a pesquisas científicas frutíferas, o pensamento de que os processos naturais podem realmente impedir a ocorrência de grandes mudanças evolutivas seria um anátema para os cientistas que defendem alguma forma de naturalismo filosófico ou religião secular.
Em uma recente conferência AAAS, Michael Ruse, filósofo da ciência e famoso defensor do darwinismo, admitiu que a teoria evolucionária se baseia em suposições metafísicas.
“E, certamente, não há dúvida sobre isso, que no passado, e eu acho que também no presente, para muitos evolucionistas, a evolução funcionou como algo com elementos que são, digamos, semelhantes a uma religião secular ... E parece-me muito claro que em algum nível muito básico, a evolução como uma teoria científica se compromete com um tipo de naturalismo, a saber, que em algum nível alguém vai excluir milagres e esse tipo de coisa aconteça o que acontecer. ” (Ruse, 1993)
Em outras palavras, não importa o que a evidência infere, não importa o quão inadequada a teoria darwiniana possa ser, apenas explicações naturalísticas para questões de origens serão consideradas científicas.
Se assumirmos que a ciência se limita ao estudo de processos puramente materialistas e, além disso, assumirmos que a origem da vida e a origem dos planos corporais principais estão sujeitos à investigação científica, então a única explicação verdadeiramente científica das origens deve ser materialista e naturalista por definição. De acordo com essas regras de raciocínio, alguma forma de darwinismo é a única explicação "científica" possível das origens. A criação em qualquer forma, incluindo evolução teísta, é por definição "não científica".
Esse raciocínio é refletido em um livreto publicado pela National Academy of Sciences intitulado Science and Creationism (1984). A influente National Academy of Sciences, que representa os cientistas mais notáveis do país, argumentou que o conceito de criação não é científico:
... falha em exibir a característica mais básica da ciência: a confiança em explicações naturalísticas. Em vez disso, os proponentes da “ciência da criação” sustentam que a criação do universo, da terra, dos seres vivos e do homem foi realizada por meios sobrenaturais inacessíveis ao entendimento humano (National Academy of Sciences, 1984).
A National Academy of Sciences simplesmente definiu todas as alternativas à evolução puramente naturalística, insistindo que apenas explicações naturalistas podem ser consideradas para responder a questões de origens últimas. Por definição, não há alternativa científica para a ideia de que “o homem é o resultado de um processo natural e sem propósito que não o tinha em mente” (Simpson, 1967).
Os defensores da “ciência da criação” ou defensores do “design inteligente” nunca pretendem explicar o mecanismo pelo qual o universo, a terra, os seres vivos ou o homem passaram a existir. Em sua maior parte, eles tentaram criticar a teoria evolucionária e apontar áreas da teoria que não podem ser testadas ou estão em conflito com os dados empíricos. Ao fazer isso, eles inferiram que os processos puramente mecanicistas são insuficientes para dar conta da ordem e da complexidade do cosmos. Isso provocou uma reação quase religiosa da Academia:
A “ciência da criação” é, portanto, manifestamente um dispositivo projetado para diluir o poder de persuasão da teoria da evolução. O modo dualista de análise e a argumentação negativa
empregado para realizar essa diluição é, além disso, antitético ao método científico (National Academy of Sciences, 1984).
O professor de direito de Berkeley, Phillip E. Johnson, em seu livro recente, Darwin On Trial , conclui:
A Academia, portanto, definiu “ciência” de tal maneira que os defensores da criação sobrenatural não podem argumentar por sua própria posição nem contestar as reivindicações do estabelecimento científico. Essa pode ser uma maneira de ganhar uma discussão, mas não é satisfatória para quem pensa ser possível que Deus realmente teve algo a ver com a criação da humanidade, ou que algumas das afirmações que os cientistas fazem sob o título de "evolução" podem ser falso (Johnson, 1991).
Os cientistas e educadores de ciências devem continuar a operar sob a suposição de que as origens devem ser explicadas por processos naturalísticos puramente não guiados? Apesar do fato de que todas as outras explicações das origens não atendem à definição de ciência da Academia, a resposta pode ser dada como um claro, Não. Dado o fato de que os eventos da criação são possibilidades históricas, é inteiramente possível que os processos naturais por si só sejam insuficientes. para explicar a origem da vida e todas as informações genéticas. Embora o naturalismo filosófico exija um cosmos sem um criador, a ciência não. Os cientistas que trabalham dentro dos limites do naturalismo metodológico poderiam, de fato, descobrir processos naturais que evitam que grandes mudanças evolutivas ocorram, processos que explicam os padrões difusos do nível de táxon estase.
Sem dúvida, os cientistas precisarão obter uma compreensão muito maior dos processos subjacentes à estase. A estase das espécies comumente continua por milhões de anos, períodos para os quais a constância ambiental não parece possível. Na verdade, a estagnação das espécies freqüentemente parece persistir, apesar das evidências de mudanças ambientais. A seleção natural é obviamente apenas uma parte de todo o quadro. Mecanismos genéticos internos e de desenvolvimento podem desempenhar um papel ainda maior na manutenção da estase, inibindo o desenvolvimento de formas transicionais.
Se Mivart estava correto ao concluir que a seleção natural é incompetente para explicar os estágios incipientes das estruturas úteis, podemos apenas criticá-lo por não levar sua ideia longe o suficiente. Ele poderia ter desenvolvido uma teoria de “macrostase” e estabelecido a seleção natural como um mecanismo-chave subjacente ao fenômeno da estabilidade morfológica e o mecanismo que explica a falta de formas de transição no registro fóssil. Se ele tivesse feito isso, poderíamos ter emergido do século XIX com duas teorias principais da história natural: uma explicando as mudanças evolutivas menores e a ancestralidade comum dos táxons inferiores e outra explicando a estabilidade dos táxons superiores. Mais importante ainda, teríamos entrado no século vinte com teorias que refletissem com mais precisão os dados empíricos.
Ao entrarmos no vigésimo primeiro século devemos fazer uma pausa e re-examinar os nossos pressupostos, bem como os nossos dados. Devemos ter cuidado para não escorregar para o cientificismo e devemos nos esforçar constantemente para descrever a natureza com mais precisão, mesmo que isso signifique descartar algumas de nossas crenças mais queridas. Como disse Pierre-Paul Grasse ', ex-presidente da Academie des Sciences francesa e editor do Traite' de Zoologie, de 35 volumes :
Hoje, nosso dever é destruir o mito da evolução, considerado um fenômeno simples, compreendido e explicado, que se desdobra rapidamente diante de nós. Os biólogos devem ser encorajados a pensar sobre as fraquezas e extrapolações que os teóricos apresentam ou definem como verdades estabelecidas. O engano às vezes é inconsciente, mas nem sempre, já que algumas pessoas, devido ao seu sectarismo, propositalmente negligenciam a realidade e se recusam a reconhecer a inadequação e falsidade de suas crenças (Grasse, 1977).
Devemos ter em mente que, apenas porque a evolução neodarwiniana é a explicação naturalista mais plausível das origens, não devemos supor que ela seja necessariamente verdadeira. Da mesma forma, apenas porque a criação envolve processos que não são naturais, não devemos assumir que os eventos da criação - sejam repentinos ou graduais - não ocorreram. Não seria razoável supor isso. Os eventos de criação podem não estar sujeitos a investigação científica, mas a estase definitivamente está. “Stasis são dados.”
Em retrospecto, parece que Darwin estava menos preocupado com a questão científica de explicar com precisão os dados empíricos da história natural e mais preocupado com a questão religiosa ou filosófica de explicar o design de todas as espécies sem um designer. A teoria geral da evolução de Darwin pode, em última análise, ser pouco mais do que uma extrapolação injustificada da microevolução baseada mais na filosofia do que nos fatos. O problema é que o darwinismo continua a distorcer a ciência do século XX.
REFERÊNCIAS
Bowring, SA, Grotzinger, JP, Isachsen, CE, Knoll, AH, Pelechaty, SM, Kolosov, P., “Calibratting Rates of Early Cambrian Evolution,” Science , vol. 261, 3 de setembro de 1993, pp. 1293-1298
Darwin, C. (1859), The Origin of Species (Reimpressão da primeira edição),
Avenel Books, Crown Publishers, Nova York, 1979
Darwin, C. (1871), The Descent of Man, and Selection in Relation to Sex,
J. Murray, Londres
Erwin, D., Valentine, J. e Sepkoski, J. (1988),
“Um Estudo Comparativo de Eventos de Diversificação”, Evolution , vol. 41, pág. 1183
Futuyma, DJ, (1986), Evolutionary Biology, Sinauer Associates Inc.,
Sunderland, MA, p. 2
Gould, SJ (1977), “Evolution's Erratic Pace”, Natural History , vol. 86, maio
Gould, SJ (1980), "É uma teoria nova e geral da evolução emergente?"
Paleobiologia, 6 (1), p. 120
Gould, SJ (1985), "Não necessariamente uma asa", História Natural , outubro, pp. 12, 13
Gould, SJ (1988), "A Web of Tales", Natural History , outubro, pp. 16-23
Gould, SJ, (1989), Wonderful Life , WW Norton and Company, Nova York, p. 49
Gould, SJ (1991), “Opus 200”, História Natural , agosto, p. 16
Grasse, Pierre-Paul (1977), Evolution of Living Organisms , Academic Press, New York, NY, p.8
Horan, P. (1979), Prefácio para The Origin of Species, Charles Darwin, 1859, (Reimpressão da primeira edição), Avenel Books, Crown Publishers, New York, p. viii
Johnson, P. (1991), Darwin on Trial , Regnery Gateway, p. 8
Kerr, R. (1993), “Evolution's Big Bang Gets Even More Explosive,” Science , vol. 261, 3 de setembro de 1993, p. 1274
Levi-Setti, R. (1975), Trilobites: A Photographic Atlas , University of Chicago Press, pp. 23-45
21
Lewin, R. (1980), “Evolutionary Theory Under Fire”, Science, vol. 210, 21 de novembro, p. 883
Lewin, R. (1988), Science , vol. 241, 15 de julho, p. 291
Lovtrup, S. (1987), Darwinism: The Refutation of a Myth,
Croom Helm Ltd., Beckingham, Kent, p. 275
Macnab, R. (1978), “Bacterial Mobility and Chemotaxis: The Molecular Biology of a Behavioral System”, CRC Critical Reviews in Biochemistry , vol. 5, edição 4, dezembro, pp. 291-341
Mivart, G. (1871), On the Genesis of Species, Londres, Macmillan
Academia Nacional de Ciências (1984),
Ciência e criacionismo: uma visão da Academia Nacional de Ciências
Raup, D. (1979), "Conflicts Between Darwin and Paleontology",
Field Museum of Natural History Bulletin , vol. 50 (1), pág. 24, 25
Raup, DM (1981), “Evolution and the Fossil Record”, Science , 17 de julho, p. 289
Ruse, M. (1993), "Nonliteralist Antievolution", Annual Meeting of the AAAS, 13 de fevereiro de 1993, no Symposium "The New Antievolutionism"
Simpson, GG (1944), Tempo and Mode in Evolution , Columbia University Press, Nova York, p. 105, 107
Simpson, GG (1967), The Meaning of Evolution, p. 345 (edição revisada)
Thompson, WR (1956), Introdução à Origem das Espécies, (Reimpressão da primeira edição), Charles Darwin, Biblioteca Everyman, no. 811, Dent
Valentine, JW, Awramik, SM, Signor, PW e Sadler, PM (1991), "The Biological Explosion at the Precambrian-Cambrian Boundary," Evolutionary Biology, Vol. 25, Max K. Hecht, editor, Plenum Press, New York and London, p.284
Valentine, J. e Erwin, D. (1985)
Desenvolvimento de “Interpretando Grandes Experimentos de Desenvolvimento: O Registro Fóssil” como um Processo Evolutivo
Rudolf A. Raff e Elizabeth C. Raff, Editores Alan R. Liss, Inc., Nova York, pp. 71, 95, 96
Wise, K. (1991), “Changing Stasis”, Origins Research , vol. 13, não. 1, pág. 20-22
Sodré GB Neto (2020) " Marcadores Biomoleculares, NGS e a Obscuridade de Diagnóstico" citando:
- ↑ Patel, Ravi K.; Jain, Mukesh (2012). «NGS QC Toolkit: A Toolkit for Quality Control of Next Generation Sequencing Data». PLOS ONE (em inglês). 7 (2): e30619. ISSN 1932-6203. PMC 3270013
 . PMID 22312429. doi:10.1371/journal.pone.0030619
. PMID 22312429. doi:10.1371/journal.pone.0030619 - ↑ Basener, William F.; Sanford, John C. (2018). «The fundamental theorem of natural selection with mutations». Journal of Mathematical Biology. 76 (7): 1589–1622. ISSN 1432-1416. PMC 5906570. PMID 29116373. doi:10.1007/s00285-017-1190-x
- ↑ Borowska, Alicja; Szwaczkowski, Tomasz; Kamiński, Stanisław; Hering, Dorota M.; Kordan, Władysław; Lecewicz, Marek (1 de maio de 2018). «Identification of genome regions determining semen quality in Holstein-Friesian bulls using information theory». Animal Reproduction Science (em inglês). 192: 206–215. ISSN 0378-4320. doi:10.1016/j.anireprosci.2018.03.012
- ↑ Casellas, J.; Varona, L. (1 de agosto de 2011). «Short communication: Effect of mutation age on genomic predictions». Journal of Dairy Science (em inglês). 94 (8): 4224–4229. ISSN 0022-0302. doi:10.3168/jds.2011-4186
- ↑ Shmookler Reis, Robert J.; Ebert, Robert H. (1 de janeiro de 1996). «Genetics of aging: Current animal models». Experimental Gerontology. In Memory of Samuel Goldstein 1938-1994 (em inglês). 31 (1): 69–81. ISSN 0531-5565. doi:10.1016/0531-5565(95)00019-4
- ↑ Crevecoeur, Guibert U. (1 de dezembro de 2019). «Entropy growth and information gain in operating organized systems». AIP Advances. 9 (12). 125041 páginas. doi:10.1063/1.5128315
- ↑ Crevecoeur, Guibert U. (1 de dezembro de 2019). «Entropy growth and information gain in operating organized systems». AIP Advances. 9 (12). 125041 páginas. ISSN 2158-3226. doi:10.1063/1.5128315
- ↑ Brewer, Wesley H.; Baumgardner, John R.; Sanford, John C. (10 de abril de 2013). «Using Numerical Simulation to Test the ?Mutation-Count? Hypothesis». WORLD SCIENTIFIC: 298–311. ISBN 978-981-4508-71-1. doi:10.1142/9789814508728_0012
- ↑ Carter, Robert W; Sanford, John C (12 de outubro de 2012). «A new look at an old virus: patterns of mutation accumulation in the human H1N1 influenza virus since 1918». Theoretical Biology & Medical Modelling. 9. ISSN 1742-4682. PMC 3507676. PMID 23062055. doi:10.1186/1742-4682-9-42







