
por Jim Gibson Geoscience Research Institute Tradução e comentários de Sodré GB Neto Link Original: https://www.grisda.org/origins-23068
RESUMO
Um dos desafios mais interessantes no entendimento da história da Terra é a explicação da ordem no registro fóssil. A identificação e análise de padrões fósseis podem fornecer uma das ferramentas necessárias para alcançar uma melhor compreensão do registro fóssil. Padrões fósseis e tendências fósseis que se estendem ao longo do registro fóssil implicam que alguns processos agiram durante toda a produção desse registro. Neste artigo, 25 padrões fósseis relatados são classificados em quatro categorias: padrões de diversidade fóssil; padrões morfológicos fósseis; padrões ecológicos fósseis; e padrões deposicionais. São possíveis possíveis interpretações criacionistas e evolucionárias desses padrões e tendências fósseis. Alguns padrões fósseis parecem difíceis de explicar do ponto de vista criacionista; outros parecem difíceis de explicar do ponto de vista evolutivo.
O estudo do registro fóssil revelou muito sobre o passado. Nosso conhecimento foi desenvolvido através do estudo de características como estruturas anatômicas, o grau de preservação, os tipos de fósseis encontrados juntos e a natureza dos sedimentos circundantes. Com o acúmulo desses dados, é natural que estudos comparativos sejam realizados para determinar quais padrões podem ser identificados. Padrões no registro fóssil podem fornecer pistas valiosas para identificar processos ativos durante a produção do registro fóssil. Este artigo tem como objetivo pesquisar e classificar os tipos de padrões fósseis relatados na literatura e comentar seu possível significado. A maioria dos padrões fósseis relatados aqui são da porção Fanerozóica da coluna geológica (veja a Figura 1), mas alguns padrões pré-cambrianos estão incluídos.


Muitos padrões gerais no registro fóssil foram relatados. Padrões fósseis que mostram uma mudança direcional sustentada são aqui denominados tendências fósseis. Podemos distinguir dois tipos de tendências fósseis. [1] Existe uma tendência de substituição de fósseis quando fósseis com certas características são substituídos por fósseis com características diferentes. Uma tendência de adição de fósseis ocorre onde fósseis com certas características são unidos por fósseis com características diferentes. Ambos os tipos de tendências são encontrados no registro fóssil. Esses diferentes tipos de tendências podem ter explicações causais diferentes; portanto, é importante observar que tipo de tendência está envolvido em um determinado padrão. Todos os padrões são generalizações e podem ocorrer exceções.
A maioria dos padrões fósseis pode ser colocada em uma das quatro categorias. (1) Padrões de diversidade são aqueles que se relacionam com as frequências de táxons fósseis. (2) Os padrões morfológicos são aqueles que se relacionam às características morfológicas dos táxons fósseis. (3) Os padrões ecológicos envolvem a consideração dos tipos de habitats representados pelos fósseis, sem preocupação com o grupo taxonômico. (4) Padrões deposicionais são aqueles que se relacionam com os tipos de sedimentos nos quais os fósseis são preservados; por exemplo, se condições de enterro catastróficas são indicadas.
Centenas de exemplos de padrões fósseis foram relatados, muito maiores do que podem ser mencionados aqui. Ao agrupá-los em categorias, é possível descrever padrões fósseis representativos e tentar uma avaliação de sua significância (ver Apêndices 1-3).
Principais padrões no registro fóssil
Padrões de diversidade
Rotatividade biótica repetida . Os padrões de diversidade formam algumas das características mais conspícuas e significativas do registro fóssil. Uma das características mais importantes do registro fóssil é a separação de diferentes tipos de fósseis em diferentes estratos. [2] (Esse padrão não aparece tão bem nos estratos pré-cambrianos. [3]) Essa separação de fósseis em diferentes camadas é tão consistente que os cientistas costumam usá-los para auxiliar na classificação dos sedimentos. Como exemplo, considere o grupo extinto de artrópodes conhecido como trilobitas. Os trilobitas são encontrados apenas em rochas paleozóicas. Certos tipos de trilobitas ocorrem apenas em sedimentos cambrianos, outros ocorrem apenas em sedimentos ordovicianos e alguns ocorrem apenas em outras camadas. Da mesma forma, os dinossauros são encontrados apenas em rochas mesozóicas, com diferentes tipos de dinossauros nas camadas Triássica, Jurássica e Cretáceo. A rotatividade de bióticos é um padrão de substituição e contém dois padrões de componentes marcantes, discutidos nas duas seções a seguir.
Os cientistas observaram com que consistência os fósseis são organizados em camadas, e organizaram essas camadas em sequência e as compararam com camadas de outras regiões. Uma espécie de "sequência principal" foi preparada. Essa sequência principal de fósseis é conhecida como coluna bioestratigráfica (veja a Figura 1). Estratos com fósseis cenozóicos ocorrem no topo da coluna, com estratos contendo fósseis mesozóicos abaixo deles e rochas contendo fósseis paleozóicos abaixo dos estratos mesozóicos. As camadas pré-cambrianas ocorrem abaixo do Paleozóico.
Aparições coordenadas . Uma característica marcante do registro fóssil é o surgimento repentino de vários tipos de fósseis em vários locais da Terra no mesmo ponto da coluna geológica. O exemplo mais famoso ocorre nas rochas cambrianas, que ficam na base das rochas paleozóicas, [4]e é chamado de "explosão cambriana". Rochas pré-cambrianas contêm relativamente poucos fósseis, a maioria dos quais parecem ser bactérias. Existem algumas impressões fósseis estranhas abaixo do cambriano, conhecidas como fósseis ediacaranos, que podem representar organismos multicelulares. Mas uma grande proporção dos principais grupos de invertebrados com partes esqueléticas duras é representada como fóssil nos estratos cambrianos (veja a Figura 2). Muitos filos de animais de corpo mole estão ausentes no registro cambriano, mas isso reflete a incompletude do registro fóssil, não a ausência desses filos durante a deposição de sedimentos cambrianos. Muitos outros exemplos de aparência coordenada ocorrem, [5]mas a explosão cambriana é de longe o exemplo mais espetacular. A aparência coordenada é um padrão de adição que persiste ao longo do registro fóssil. Nenhuma tendência foi relatada para esse padrão.
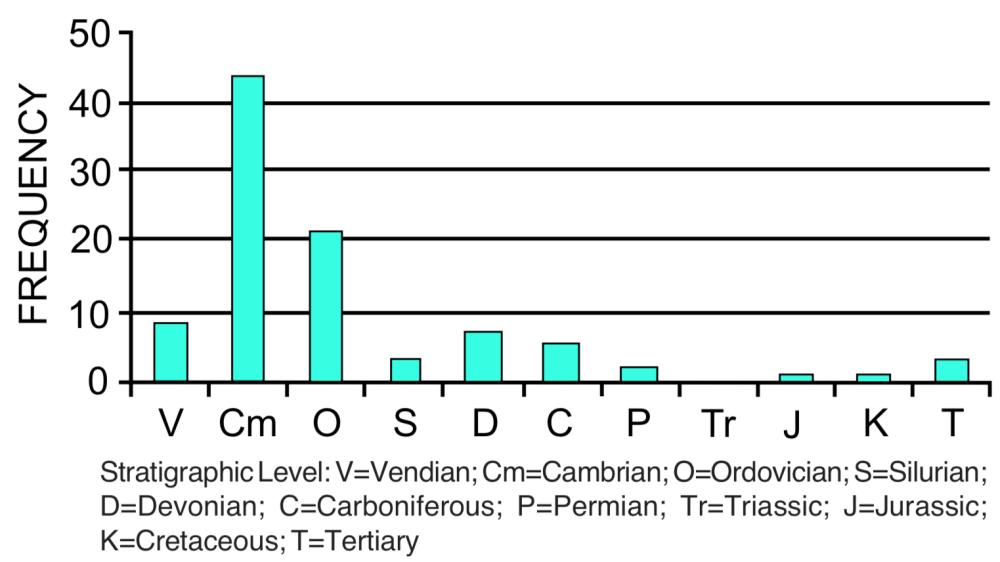
Desaparecimentos coordenados . Um grande número de espécies fósseis pode desaparecer do registro geológico em um nível estratigráfico específico (veja a Figura 3). O desaparecimento nunca está completo, mas existem vários exemplos em que as estimativas indicam que mais de 50% das espécies desaparecem no mesmo nível estratigráfico. [6]As fronteiras entre os níveis estratigráficos são frequentemente identificadas com base em desaparecimentos coordenados. O maior exemplo disso é o desaparecimento de quase metade das famílias (ver Figura 3) e cerca de 95% de todas as espécies no topo do Paleozóico. Dinossauros e muitos outros grupos de répteis e invertebrados marinhos desaparecem do registro no topo do Mesozóico. Outros exemplos de desaparecimentos coordenados em grande escala ocorrem no topo do Ordoviciano, próximo ao Devoniano e ao Triássico. O desaparecimento coordenado é um padrão de subtração. Nenhuma tendência sustentada nesse padrão foi relatada.

Diversidade crescente . O número de espécies geralmente aumenta à medida que a pessoa se move para cima através do registro fóssil. O aumento é altamente irregular, mas a tendência geral é clara. [7]Por exemplo, o número de espécies conhecidas das rochas cambrianas é de aproximadamente 8.000, aumentando para 15.000 no Carbonífero (Paleozóico superior). A diversidade total de espécies diminui, mas aumenta para 22.000 em rochas cretáceas e 43.000 em rochas cenozóicas. Tendências semelhantes em relação ao aumento da diversidade são observadas para gêneros e famílias (veja a Figura 3), mas não para filos e classes (veja a próxima seção). A diversidade também aumenta frequentemente dentro de um táxon; por exemplo, o número de espécies ou gêneros pode aumentar em uma família ou categoria superior. O aumento da diversidade é uma tendência que envolve adição e substituição, sendo a adição dominante. Uma exceção notável a esse padrão é o aumento e a diminuição da diversidade de microfósseis nas rochas pré-cambrianas. [8]
Disparidade antes da diversidade . Disparidade refere-se à extensão da divergência morfológica entre os membros de um grupo, enquanto diversidade refere-se ao número de táxons dentro de um grupo. Surpreendentemente, o número de espécies fósseis (diversidade) no Cambriano é baixo, mas o número de filos e classes (disparidade) é alto, em comparação com o número em outras partes da coluna geológica. Em geral, cada filo ou classe de fósseis cambrianos contém apenas algumas espécies, enquanto esses mesmos grupos podem ter um número maior de espécies nos estratos acima do cambriano. Os estratos acima do Cambriano contêm um número maior de espécies e famílias, mas poucos filos adicionais. Assim, os fósseis cambrianos são altamente díspares, mas o número de espécies (diversidade) é relativamente baixo. Esse padrão foi chamado de "disparidade antes da diversidade"[9] Existem outros exemplos na coluna geológica onde a disparidade precede a diversidade dentro de certos grupos, [10] mas a Explosão Cambriana é o maior exemplo. Esse padrão possui apenas um componente direcional muito fraco e provavelmente não se qualifica como uma tendência fóssil.
Provincialidade . A província refere-se à distinção entre assembléias fósseis de diferentes regiões geográficas. Diz-se que a província é alta quando cada região tem um conjunto fóssil distinto e baixa quando o número de conjuntos fósseis distintos é baixo. Os padrões estratigráficos da província geralmente requerem mais dados do que os que estão prontamente disponíveis. No entanto, tem sido relatado que a província de biotas terrestres tende a aumentar através do registro fóssil. [11]
Padrões Morfológicos
Aumento da complexidade . As tendências morfológicas mais intimamente relacionadas à teoria da ancestralidade comum são de especial importância para as discussões sobre criação e evolução. Uma das tendências mais amplamente relatadas é o aumento da complexidade, de bactérias nas rochas pré-cambrianas a seres humanos nos estratos cenozóicos. Às vezes, isso é visto como o principal tema da evolução, do simples ao complexo. Complexidade é um conceito difícil de quantificar, mas o número de tipos de células tem sido usado como um estimador de complexidade. [12]No entanto, os dados do tipo de célula que suportam essa tendência parecem correlacionados com a sequência de vertebrados (ver Figura 4) e podem ser um subproduto acidental dessa sequência. A tendência para aumentar a complexidade é na verdade uma tendência de adição, não uma tendência de substituição. Não há evidências de que as bactérias vivas sejam mais complexas que as encontradas nas rochas pré-cambrianas. [13] A tendência para aumentar a complexidade está correlacionada com a tendência para aumentar a diversidade. Certos grupos considerados mais complexos, particularmente grupos de vertebrados e plantas, aparecem progressivamente no registro geológico em uma sequência que corresponde à crescente complexidade.

Espécie morfológica-estase . Estase morfológica é a persistência da morfologia através de partes da coluna geológica. [14] Embora haja alguma disputa sobre esse padrão, parece que a maioria dos paleontologistas aceita a predominância de estase morfológica nas espécies. Uma espécie fóssil normalmente parece a mesma na primeira e na última aparição. Espécimes individuais podem mostrar pequenas variações em torno de alguma média, mas geralmente não há direcionamento para diferenças morfológicas dentro de uma espécie. Inúmeros exemplos de mudança direcional foram propostos, [15] mas afirma-se que representam uma minoria de casos, e alguns foram reinterpretados por outros estudos. [16]O caso mais extremo de estase são provavelmente as cianobactérias, que parecem as mesmas nos sedimentos pré-cambrianos e nas populações modernas. [17]
Estase morfológica de táxon superior . A estase morfológica em categorias taxonômicas mais altas [18] refere-se à persistência de planos corporais em categorias taxonômicas mais altas que as espécies. Por exemplo, muitos planos corporais de invertebrados nos níveis de filo e classe persistem por todo o Fanerozóico. Essa persistência não produz uma tendência fóssil, mas continua ao longo do registro fóssil. A estase de táxons mais altos está relacionada à aparência de disparidade antes da diversidade, discutida acima.
Estase coordenada . Estase coordenada refere-se à observação de que grupos de espécies em uma formação geológica específica, ou parte de uma formação, podem permanecer essencialmente inalterados por sedimentos que são interpretados como representando milhões de anos de tempo. [19] Fósseis exibindo estase coordenada podem ocorrer em sedimentos delimitados acima e abaixo por horizontes de alta rotatividade biótica. Atualmente, esse padrão é controverso e são necessários mais estudos para testar sua significância. Nenhuma direção foi relatada para este padrão.
Lacunas morfológicas entre as espécies . As espécies fósseis são tipicamente separadas umas das outras por lacunas na morfologia. [20]Isso causa o aparecimento abrupto típico de espécies fósseis. Obviamente, deve-se lembrar que as espécies fósseis são tipicamente identificadas com base na morfologia. É a existência de lacunas morfológicas que permite distinguir diferentes espécies fósseis. Duas espécies fósseis que se classificam umas nas outras podem ser reconhecidas como uma única espécie com variabilidade maior que a média. São conhecidas espécies fósseis com variabilidade acima da média, mas essa situação também pode ser encontrada em algumas espécies vivas. Em termos gerais, parece que a variação individual dentro das espécies fósseis é geralmente da mesma magnitude que nas espécies vivas. Esse padrão persiste ao longo do registro fóssil, sem nenhuma tendência direcional.
Lacunas morfológicas formam um padrão hierárquico aninhado . As morfologias dos fósseis geralmente podem ser organizadas para formar um padrão hierarquicamente aninhado, formando a base do atual sistema de categorias taxonômicas. Um grupo de espécies separadas por pequenas lacunas morfológicas compreende um gênero. Os gêneros são separados por lacunas morfológicas maiores. As diferenças são cada vez maiores, considerando-se categorias taxonômicas mais altas, como famílias, ordens e classes. [21] Novas descobertas fósseis às vezes reduzem o tamanho das lacunas, especialmente em níveis taxonômicos mais baixos, [22]mas as lacunas nos níveis taxonômicos mais altos são notavelmente distintas. Esse padrão não parece resultar em nenhuma tendência direcional através do registro fóssil. Exceções ocasionais a esse padrão ocorrem na forma de "mosaicos morfológicos" espécies com uma mistura de características de dois ou mais grupos morfologicamente distintos. Tais espécies podem indicar a natureza artificial do nosso sistema taxonômico.
Mudanças no tamanho do corpo . O tamanho do corpo geralmente mostra uma tendência direcional para as espécies dentro de um grupo. [23] As tendências para aumentar o tamanho são mais conhecidas, mas também são relatadas tendências para diminuir o tamanho. A maioria das tendências entre os tetrápodes (anfíbios, répteis, aves e mamíferos) envolve tamanho. [24] As tendências no tamanho do corpo podem ser tendências de adição ou tendências de substituição. As tendências no tamanho do corpo são estratigraficamente limitadas, geralmente se estendem por apenas uma ou algumas divisões estratigráficas.
Séries morfológicas . As espécies fósseis podem frequentemente ser organizadas em uma série morfológica na qual a direcionalidade da mudança morfológica é consistente com a sequência estratigráfica dos fósseis. [25] O exemplo mais famoso disso é a série de cavalos, que começa com uma espécie de 5 dedos (que pode ou não ser um cavalo) no Eoceno (terciário inferior) e progride para um grupo de espécies com 3 dedos. o Oligoceno e o Mioceno (terciário médio) e termina com cavalos vivos de um dedo. Esta tendência é acompanhada por uma tendência ao aumento do tamanho corporal. [26] Outra série morfológica são as crescentes características semelhantes a mamíferos nos répteis sinapsídeos das rochas do Paleozóico superior e do Mesozóico inferior. [27]Outros exemplos incluem a série de dinossauros a pássaros [28], de mamíferos terrestres a baleias [29] e entre grupos de invertebrados. [30] As séries morfológicas são principalmente tendências de substituição, mas também mais podem ser algumas adições.
Modernidade crescente . A maioria dos fósseis é de espécies extintas, mas alguns são mais semelhantes às espécies vivas do que outros. Os fósseis do alto da coluna estratigráfica se assemelham mais às espécies vivas do que os fósseis do baixo da coluna. [31]Por exemplo, os moluscos cenozóicos são bastante semelhantes às espécies vivas, enquanto os moluscos mesozóicos são menos semelhantes, e os moluscos paleozóicos são bem diferentes dos que vivem atualmente (Figura 5). Essa tendência é vista também entre os vertebrados. Os peixes paleozóicos são principalmente peixes de aparência estranha, diferente de qualquer outro ser vivo atualmente. Os peixes mesozóicos são mais semelhantes aos peixes vivos, e os peixes cenozóicos são bastante semelhantes aos tipos vivos de peixes. Essa tendência de substituição é bem conhecida e se aplica a quase todos os grupos de organismos, exceto as bactérias, que parecem ter mudado muito pouco. [32]
Especialização onipresente . Especialização de uma espécie significa que a espécie possui estruturas morfológicas que parecem apropriadas para habitats específicos ou papéis ecológicos. Praticamente todas as espécies fósseis são especializadas de alguma forma. Isso é ilustrado pelos artrópodes da Explosão Cambriana, como apontado por Gould. [33] Diz-se que espécies sem especialização notável são generalizadas. A maioria das espécies possui algumas características relativamente generalizadas, enquanto outras podem ser altamente especializadas. Esse é um padrão geral no registro fóssil e não forma uma tendência direcional.

Falta de antepassados identificáveis . O registro fóssil contém mais de duzentas mil espécies. Encontrar relações entre essas espécies é problemático. [34]Os táxons mais altos são freqüentemente chamados de ancestrais a outros táxons mais altos, mas os evolucionistas reconhecem que os táxons mais altos não podem ser ancestrais reais de nada, pois são construções taxonômicas e não entidades reais. Grupos de espécies com diferenças morfológicas sucessivamente menores podem ser identificados e relações genealógicas podem ser propostas. No entanto, é notavelmente difícil identificar uma espécie fóssil como diretamente ancestral de outra. A dificuldade é agravada à medida que a categoria taxonômica em discussão aumenta. Uma das principais razões para a dificuldade é que quase todas as espécies têm alguma especialização que as impede de ascendência direta de qualquer outra espécie conhecida. Nenhuma direcionalidade para esse padrão foi relatada.
Padrões Ecológicos
Aumento da diversidade de habitats . O número de habitats representados aumenta à medida que se move para cima através da coluna geológica. [35]Rochas pré-cambrianas são dominadas por fósseis de bactérias. As rochas cambrianas possuem apenas fósseis marinhos, principalmente de espécies que vivem em substratos duros no fundo do mar. Fósseis de espécies de água doce aparecem pela primeira vez em números no Siluriano (Paleozóico médio), embora existam algumas espécies possíveis de água doce em depósitos mais baixos. Espécies totalmente terrestres são relatadas a partir de rochas silurianas, mas estão melhor representadas em rochas devonianas. Rochas mesozóicas contêm fósseis de uma maior diversidade de habitats, e rochas cenozóicas continuam a tendência de maior diversidade de habitats representados. Este princípio também se estende à ecologia em menor escala. Vinte guildas ecológicas foram identificadas no reino marinho. [36]Nove deles estão presentes nos cambrianos, 14 estão presentes no paleozóico como um todo e 20 estão presentes nos fósseis cenozóicos. Este é outro exemplo de tendência por adição, neste caso uma "expansão ecológica".
Aumento da terrestre . Algumas tendências em direção ao aumento da terrestre no registro fóssil foram relatadas. Fósseis dos estratos paleozóicos mais baixos são todas criaturas marinhas que aparentemente viveram no fundo ou próximas ao fundo do mar. [37] No Paleozóico médio, encontram-se não apenas criaturas marinhas, mas também muitos fósseis de espécies que aparentemente viviam em pântanos ou talvez ao longo das margens dos mares ou rios. No Paleozóico superior, encontram-se espécies totalmente terrestres. Os fósseis mesozóico e cenozóico incluem representantes de todos os habitats ecológicos anteriores. Os exemplos mais claros dessa tendência são dados pela sequência das primeiras aparições de grupos de plantas [38] e vertebrados. [39]Os vertebrados mais baixos da coluna geológica são os peixes, que requerem água. O próximo tipo ecológico a aparecer são os anfíbios, que vivem ao longo das margens da água. No alto da coluna, eles se juntam aos répteis, que podem viver longe da água. Mamíferos e pássaros são as últimas classes de vertebrados a aparecerem como fósseis. O aumento da terrestre não é uma tendência forte, porque ocorrem exceções. A exceção mais notável pode ser a predominância de (provavelmente) bactérias fotossintéticas no pré-cambriano, embora tenha havido sugestões de que possam ter sido contaminantes do subsolo. [40] É importante observar que essa tendência não é uma tendência de substituição, mas uma tendência de adição, porque todos esses habitats ainda estão ocupados.
Maior mobilidade . Uma possível tendência para o aumento da mobilidade foi relatada entre os invertebrados marinhos. [41] Diz-se que fósseis paleozóicos são dominados por espécies que vivem no fundo do mar ou próximas a ele e com mobilidade limitada. Fósseis marinhos mesozóicos e cenozóicos tendem a incluir mais tipos móveis. Em terra, pode haver um efeito semelhante produzido pelas tendências de aumento de tamanho, como parece ser o caso de alguns dos dinossauros e mamíferos. [42] Não é certo o quão comum é essa tendência. Se a tendência for válida, seria outra tendência por adição.
Padrões em ambientes deposicionais
Depósitos de tempestade . Informações sobre padrões em ambientes deposicionais não estão tão prontamente disponíveis quanto aos outros padrões incluídos neste estudo, mas algum trabalho está sendo feito nessa área. [43] Por exemplo, relata-se que os depósitos de tempestades ocorrem com mais freqüência nas rochas ordovicianas, silurianas, devonianas, jurássicas e cretáceas (ver Figura 6). Depósitos interpretados como furacões são mais frequentes em Ordovicianos e Devonianos, enquanto tempestades de inverno inferidas são mais comuns em rochas silurianas e cretáceas.
Faunas de corpo mole bem preservadas . A maioria dos fósseis são restos de organismos de corpo duro, especialmente moluscos, equinodermes, artrópodes, vertebrados e plantas. No entanto, algumas áreas (chamadas "lagerstätten") são conhecidas pela preservação excepcional da fauna de corpo mole, incluindo minhocas, etc. [44] Essas faunas excepcionais estão espalhadas pela coluna geológica, mas podem ser super-representadas nas rochas cambrianas e jurássicas ( veja a Figura 6).

Energia deposicional para as primeiras aparições . Os sedimentos podem ser identificados como alta ou baixa energia, com base no tamanho das partículas. Partículas grandes requerem mais energia para seu transporte e deposição do que partículas pequenas. Depósitos de baixa energia, como folhelhos marinhos, são frequentemente associados à deposição em águas profundas, enquanto depósitos de energia mais altos, como arenitos marinhos, podem ser interpretados como depósitos próximos à costa. Observou-se que a maioria dos taxa mais altos de invertebrados marinhos aparece pela primeira vez em depósitos de alta energia, enquanto as últimas aparências tendem a estar em depósitos de baixa energia. [45] Essa tendência é normalmente chamada de hipótese onshore-offshore.
Ambientes deposicionais . Certos tipos de depósitos sedimentares mostram padrões ou tendências de frequência na coluna geológica. Por exemplo, 85% dos depósitos de lago inferidos ocorrem em rochas cenozóicas, 11% em rochas mesozóicas e apenas 4% em rochas paleozóicas. [46] Por outro lado, o calcário compreende uma proporção maior de rochas paleozóicas e mesozóicas, com proporções mais baixas nas rochas cenozóicas. [47]
Modos de preservação . Esta é uma área que não recebeu estudos suficientes. Alguns relatórios publicados indicam diferenças estratigráficas nos modos de preservação. [48] Por exemplo, a silicificação é declaradamente mais comum entre fósseis paleozóicos do que entre fósseis mesozóicos ou cenozóicos. São necessárias mais informações sobre esse tipo de padrão.
UMA NOTA SOBRE PADRÕES GEOGRÁFICOS
Vários padrões que refletem variação geográfica dentro de uma divisão estratigráfica foram observados no registro fóssil. [49] As tendências geográficas incluem gradientes de diversidade e variação no comprimento da faixa estratigráfica. A latitude é um fator bem conhecido que afeta as tendências geográficas. Os padrões geográficos estão além do escopo deste artigo, a menos que sejam comparados através da coluna estratigráfica.
PROBLEMAS NA INTERPRETAÇÃO DE PADRÕES E TENDÊNCIAS FÓSSEIS
Pode ser bastante difícil determinar uma causa por trás de uma tendência fóssil. De fato, tendências aparentes podem ocorrer em dados aleatórios. [50] As tendências também podem ser "caronas" que são apenas o resultado de uma tendência em algum outro recurso. [51] Por exemplo, muitas "tendências" em caracteres morfológicos estão correlacionadas com tendências em tamanho corporal ou ecologia. [52] Essas precauções devem ser lembradas ao interpretar tendências fósseis.
EXPLICANDO OS RECURSOS DO REGISTRO FÓSSEIL COMO RESULTADO DA EVOLUÇÃO
A maioria dos cientistas interpreta o registro fóssil como um registro da história evolutiva. [53]Eles explicam a segregação de fósseis em vários estratos, como resultado de mudanças que ocorrem por longos períodos de tempo. Diferentes tipos de organismos viveram em momentos diferentes e foram fossilizados quando as camadas foram depositadas em sequência. Pensa-se que as espécies que ocorrem nas rochas mais baixas sejam os ancestrais evolutivos das mais altas na coluna estratigráfica. À medida que alguém se move para cima através da coluna geológica, está se aproximando do tempo presente. Assim, deve-se esperar descobrir que os fósseis nos estratos superiores pareceriam familiares, porque estão mais intimamente relacionados às espécies vivas. Os fósseis nos estratos mais baixos estão apenas distantemente relacionados com espécies vivas ou de grupos que não existem mais. Pode-se esperar que pareçam diferentes de qualquer coisa que esteja vivendo agora. Espécies com combinações estranhas de características podem representar os tipos de estágios de transição que ocorreram à medida que novos tipos de espécies evoluíram a partir de espécies mais antigas. A expansão ecológica vista na coluna geológica reflete o fato de que a vida começou no mar. Os organismos vivos não foram capazes de viver em terra até terem desenvolvido as estruturas necessárias para sobreviver fora da água. Os evolucionistas acreditam que a teoria da evolução fornece uma boa explicação para as principais características do registro fóssil, incluindo rotatividade biótica, modernidade crescente, séries morfológicas e expansão ecológica. Os organismos vivos não foram capazes de viver em terra até terem desenvolvido as estruturas necessárias para sobreviver fora da água. Os evolucionistas acreditam que a teoria da evolução fornece uma boa explicação para as principais características do registro fóssil, incluindo rotatividade biótica, modernidade crescente, séries morfológicas e expansão ecológica. Os organismos vivos não foram capazes de viver em terra até terem desenvolvido as estruturas necessárias para sobreviver fora da água. Os evolucionistas acreditam que a teoria da evolução fornece uma boa explicação para as principais características do registro fóssil, incluindo rotatividade biótica, modernidade crescente, séries morfológicas e expansão ecológica.
Várias características do registro fóssil são pelo menos consistentes com a teoria da evolução. Uma diversidade crescente seria esperada se um único ancestral comum diversificasse e produzisse descendentes cada vez mais diversos e díspares. A evolução das adaptações para a terrestre exigiria tempo, durante o qual uma tendência para o aumento da terrestre poderia ser esperada. O aumento da complexidade e da mobilidade pode resultar da competição contínua e da expansão da ocupação ecológica. As tendências de tamanho corporal também podem resultar do aumento dos níveis de competição. [54] O aumento da concorrência também pode levar grupos marinhos mais antigos dos habitats onshore para offshore, à medida que novos grupos onshore evoluem. Espera-se um aumento na província, pois uma única massa terrestre, a Pangea, se separou e formou regiões separadas e cada vez mais isoladas.
No entanto, existem outras considerações. A teoria da evolução não fornece uma explicação tão boa para a "Explosão Cambriana". Não se esperaria que a evolução produzisse um aumento repentino de disparidade, especialmente quando se considera as grandes diferenças entre os grupos de fósseis cambrianos. A falta de ancestrais pré-cambrianos para os grupos cambrianos é outro ponto que não é facilmente explicado pela teoria da evolução, embora muitas hipóteses tenham sido propostas. [55]A estase coordenada, se válida, parece altamente anômala para a teoria da evolução. O desaparecimento coordenado, comumente chamado de "extinção em massa", parece difícil de explicar sem uma grande catástrofe global. As tendências em ambientes deposicionais também parecem sugerir vínculos sucessivos em um processo geral, em vez de uma sequência estocástica de eventos não relacionados.
A evolução também não fornece uma explicação muito boa para o padrão de lacunas morfológicas que separam os diferentes táxons mais altos de fósseis. A explicação padrão é que o registro fóssil está incompleto. As lacunas entre os taxa fósseis representam espécies extintas que realmente viveram, mas não foram descobertas como fósseis. Esses fósseis desaparecidos foram chamados de "elos perdidos". O registro fóssil certamente está incompleto, mas isso realmente explica o padrão dos fósseis? [56]As ovelhas e as vacas são morfologicamente semelhantes. As chances de encontrar mais espécies como elas parecem improváveis se o registro fóssil for altamente incompleto. Mas uma ovelha e um macaco são muito diferentes. Deve haver muitas espécies fósseis mostrando os estágios evolutivos entre uma ovelha e um macaco. Mas o oposto é verdadeiro. Existem muitos tipos de fósseis que são semelhantes aos ovinos e bovinos, respectivamente, mas os fósseis intermediários entre ovelhas e macacos estão praticamente ausentes. Se considerarmos o registro fóssil pelo valor de face, esses supostos estágios intermediários talvez nunca tenham existido. Um registro fóssil incompleto pode explicar as lacunas entre espécies próximas, mas não o padrão de lacunas entre as categorias mais altas. A especialização onipresente e a falta de ancestrais identificáveis também parecem difíceis de explicar para a teoria da evolução.
Até séries morfológicas podem ser problemáticas para a teoria da evolução. Os efeitos morfológicos observados devido à seleção natural ocorrem muito mais rapidamente do que as alterações normalmente vistas em séries fósseis. [57] Que tipo de força seletiva poderia persistir por milhões de anos, conduzindo continuamente mudanças morfológicas em pequenos incrementos? A teoria da evolução não fornece uma boa explicação para a explosão cambriana, estase coordenada, falta geral de ancestrais identificáveis ou lacunas sistemáticas entre espécies e grupos de espécies. As tendências em ambientes deposicionais também parecem pouco explicadas pela teoria da evolução. Há uma base empírica razoável para procurar outra teoria para explicar o registro fóssil.
EXPLICANDO AS CARACTERÍSTICAS DO REGISTRO FÓSSIL NO CONTEXTO DAS ESCRITURAS
Vários cientistas tentaram desenvolver interpretações criacionistas do registro fóssil, [58] com sucesso misto. Muitas das explicações são pontuais , e é necessário muito mais trabalho. No entanto, um bom começo foi feito e mais progressos podem ser esperados. Idéias de várias fontes foram incorporadas na discussão abaixo.
Algumas características do registro fóssil são prontamente explicadas do ponto de vista criacionista. A "explosão cambriana" pode ser facilmente explicada como resultado do enterro do fundo do mar nos estágios iniciais do dilúvio bíblico. [59] Os fósseis cambrianos não são relacionados entre si genealogicamente. Em vez disso, eles estão relacionados ecologicamente. São todas criaturas do fundo do mar. Outros exemplos de "aparências coordenadas" podem resultar da invasão de inundações em novas biozonas. [60] A combinação de "Explosão cambriana" e "estase com táxons mais altos" produziria o padrão de "disparidade diante da diversidade".
À medida que novas comunidades fossem encontradas pelo aumento das águas das enchentes, novos grupos de espécies seriam adicionados ao registro fóssil. Os novos grupos de espécies mostrariam apenas variação intra-específica normal, produzindo um registro de "estase coordenada". Os "desaparecimentos coordenados" ocorreriam quando uma área de origem específica estivesse esgotada ou devido a alguma mudança crítica nas condições de inundação. [61] A natureza global de muitos desaparecimentos coordenados indica um processo global. Tal "inundação em expansão" naturalmente produziria um aumento na diversidade através do registro fóssil.
A teoria da criação especial também pode explicar as lacunas morfológicas que separam os fósseis em diferentes grupos. As principais lacunas morfológicas distinguem diferentes grupos que foram criados separadamente, produzindo "estase com táxons mais altos". [62] A estase morfológica de táxons mais altos pode refletir a incapacidade de tipos criados originalmente variar naturalmente além de certos limites determinados por suas composições genéticas. A criação especial também explica a falta de ancestrais identificáveis no registro fóssil, bem como a especialização vista em praticamente todas as formas de vida.
As espécies fósseis morfológicas próximas podem representar diferenças que se acumularam em populações isoladas após a criação. A estase morfológica em nível de espécie e a notável escassez de fósseis em transição em baixos níveis taxonômicos podem ser o resultado de preservação catastrófica; não é necessário supor que nunca existiram intermediários entre espécies e gêneros similares ou que as espécies são tão estáveis quanto sugerido pelo registro fóssil. [63] Por outro lado, séries morfológicas interespecíficas podem representar triagem hidrodinâmica [64] ou clines geográficos pré-diluvianos, como são bem conhecidos na ecologia atual. [65] No caso de espécies com expectativa de vida de algumas semanas ou menos, uma tendência pode refletir uma série real de eventos de especiação.[66] Esses fatores também podem ser responsáveis por alguns dos muitos exemplos relatados de tendências no tamanho corporal.
O componente ecológico na coluna geológica também parece consistente com as teorias criacionistas, embora os detalhes não sejam bem compreendidos. Pode ser o resultado da expansão da atividade do Dilúvio. Quando o Dilúvio começou, os sedimentos seriam transportados para as elevações mais baixas primeiro. O fundo do mar provavelmente seria o primeiro a ser coberto, enterrando os organismos que ali viviam. [67] À medida que as águas subiam, grupos adicionais de organismos seriam adicionados. Eventualmente, os habitats de terras altas seriam inundados e as espécies de terras altas adicionadas à pilha de camadas fósseis, produzindo a tendência de aumento da terrestre. A mobilidade também pode ser um fator importante. [68]Esse processo também pode explicar a sequência das primeiras aparições das classes de vertebrados, da qual depende o padrão de crescente complexidade (veja a Figura 4).
Na realidade, o processo de produção do registro fóssil era muito mais complexo do que simples diferenças ecológicas. Grupos marinhos adicionais são encontrados em toda a coluna geológica, o que sugere a existência de mares continentais em diferentes elevações e em diferentes regiões geográficas. [69] Além disso, fósseis de rochas pré-cambrianas são dominados por bactérias aparentemente fotossintéticas [70] , que de alguma forma devem ter penetrado nas rochas, talvez muito cedo no Dilúvio [71] ou antes do Dilúvio [72] ou como contaminantes, [73] ou talvez presos acidentalmente em algum sistema subterrâneo envolvido no ciclo da água antes do dilúvio.
Tendências nos padrões de deposição podem refletir os estágios avançados do Dilúvio. A distribuição possivelmente não aleatória relatada de depósitos de tempestades e lagerstätten pode indicar um processo não uniforme responsável pelo registro fóssil. Uma inundação mundial pode fornecer uma explicação para isso; nesse caso, os detalhes ainda precisam ser resolvidos. Tendências deposicionais, como diminuição da deposição de calcário e aumento dos depósitos dos lagos, parecem resultados plausíveis de uma inundação mundial. As primeiras aparições dos principais táxons em depósitos interpretados como depósitos onshore podem ser o resultado da interpretação de depósitos de alta energia como onshore. Talvez os depósitos de alta energia sejam mais propensos do que os depósitos de baixa energia a capturar e preservar tipos de fósseis anteriormente não registrados.
A existência de tantos fósseis e sua preservação espetacular [74] indicam circunstâncias especiais raras no mundo de hoje. Evidências de enterro rápido, atividade geológica generalizada e atividade química de água altamente mineralizada são efeitos esperados de uma inundação mundial.
No entanto, existem algumas características da coluna geológica que os criacionistas têm mais dificuldade em explicar. A maneira consistente pela qual os fósseis são segregados na coluna geológica é uma dessas características. Alguém poderia pensar que uma inundação mundial produziria uma extensa mistura de vários tipos de fósseis. Talvez o mundo pré-diluviano tenha sido altamente estruturado, tanto ecologicamente quanto taxonomicamente. Assim, à medida que as águas do Dilúvio subiam verticalmente e se expandiam geograficamente, diferentes habitats eram engolidos e diferentes grupos taxonômicos eram sucessivamente depositados como fósseis. Isso resultaria em diferenças visíveis nos tipos de fósseis encontrados quando se compara diferentes estratos na coluna geológica.
Outra tendência difícil de explicar é a crescente semelhança com as formas modernas, quando se vê a coluna fóssil de baixo para cima. Essa tendência fóssil pode ser o resultado do alto grau de estrutura postulado para o mundo pré-diluviano. Os habitats mais próximos do fundo foram os primeiros a serem enterrados e sofreram a maior extensão de extinção. Os últimos grupos de organismos a serem engolidos pelo Dilúvio teriam as melhores chances de sobrevivência. [75] O resultado seria que as camadas inferiores teriam fósseis de espécies desconhecidas para nós agora, enquanto os estratos mais altos teriam muitos fósseis de tipos familiares de organismos.
O padrão observado também se aplica aos vertebrados terrestres, que se acredita serem preservados apenas na arca. Não está claro por que os vertebrados terrestres mostram o mesmo padrão que os invertebrados marinhos. Uma explicação sugerida para isso [76] é que as espécies de terras altas foram melhor adaptadas aos climas mais frios (e provavelmente mais severos) após o dilúvio. Outra sugestão [77] é que a sobrevivência das espécies preservadas na arca dependia da sobrevivência de espécies que não estão na arca, e aquelas espécies cujo suprimento de comida foi destruído não poderiam sobreviver.
Certas séries morfológicas também são difíceis de explicar. Os répteis sinapsídeos são um exemplo. Características crescentes semelhantes a mamíferos são vistas em fósseis sinapsídeos através do Permiano e Triássico. Um criacionista pode procurar descobrir se há alguma outra tendência associada, como uma relacionada à ecologia, [78]comportamento ou distribuição, responsável pelas séries morfológicas. É significativo que nenhuma série de ancestrais e descendentes reais possa ser identificada entre os fósseis de répteis sinapsídeos, apenas uma tendência geral para o aumento de características semelhantes a mamíferos. Embora os criacionistas tenham apontado as dificuldades de interpretar essa série como uma sequência evolucionária, eles não desenvolveram uma explicação alternativa detalhada. Foi sugerido que os répteis sinapsídeos refletem uma diversidade mais rica antes do dilúvio, [79] e que a tendência para o aumento de caracteres de mamíferos pode ser um efeito colateral de um padrão ecológico. [80]
Uma dificuldade final é a crescente província nas faunas terrestres mesozóicas e cenozóicas. Seria de esperar um aumento da província após o dilúvio, mas não está tão claro por que a província aumentaria em sedimentos que se acredita serem depositados durante o dilúvio. A província é baixa no mesozóico inferior, depois aumenta no mesozóico superior, com aumentos ainda maiores no cenozóico inferior. Esse padrão pode ser parcialmente um artefato de amostragem incompleta do registro fóssil, ou talvez seja um reflexo das diferenças biogeográficas pré-diluvianas, como entre os hemisférios norte e sul. Também poderia ser usado para indicar que o Cenozóico é um registro de repovoamento pós-diluviano [81], mas há razões para restringir o repovoamento pós-diluviano ao cenozóico superior. [82] Qualquer interpretação envolve problemas não resolvidos.
DISCUSSÃO
Aqueles que esperavam que padrões e tendências fósseis revelassem uma história direta da história da Terra encontraram desapontamento. Algumas características do registro fóssil parecem sugerir uma visão, enquanto outras parecem sugerir outra. Pessoas com visões diferentes da história da Terra podem apontar para características selecionadas do registro fóssil para apoiar suas visões.
Independentemente do ponto de vista, nosso entendimento é incompleto. O desafio para os criacionistas é explicar as tendências fósseis como resultado do modo como o dilúvio corroeu e enterrou a biota de vários habitats. O ponto de vista criacionista considera primárias as tendências ecológicas e deposicionais. Tendências de diversidade e séries morfológicas são consideradas conseqüências secundárias das tendências primárias.
Desse ponto de vista, as tendências fósseis ecológicas são interpretadas para refletir a expansão da atividade de inundação à medida que outros habitats e regiões geográficas adicionais foram varridas. Isso implica a segregação de habitats em uma ecologia pré-inundação altamente estruturada. As tendências fósseis ecológicas observadas são tendências por adição, não por substituição (consulte o Apêndice 3). Como invertebrados marinhos relativamente densos e imóveis são encontrados nas camadas superiores e inferiores, deve haver áreas de fontes adicionais disponíveis para destruição em diferentes estágios do Dilúvio. É por isso que postula-se que os habitats marinhos pré-diluvianos ocorreram em diferentes regiões e em diferentes elevações. Esta parte da explicação parece ad hoc, mas uma ecologia altamente estruturada antes do dilúvio parece ser uma parte essencial da teoria.
As tendências deposicionais são relatadas com menos frequência, mas algumas foram identificadas (consulte o Apêndice 3). A diminuição da importância relativa do calcário, juntamente com o aumento dos depósitos dos lagos, pode estar ligada à expansão do Dilúvio além dos principais corpos oceânicos no ambiente terrestre. As inferências de habitat baseadas na energia deposicional podem, alternativamente, ser consideradas sob padrões deposicionais. Mais estudos nesta área são altamente desejáveis. As localizações geográficas relativas e as posições estratigráficas dos depósitos de alta energia e baixa energia podem fornecer informações úteis sobre a sequência e extensão de vários eventos locais ou regionais durante o Dilúvio. O mesmo poderia ser dito sobre os padrões de depósitos de tempestades e lagerstätten. Seria interessante determinar se esses padrões poderiam estar relacionados a impactos extraterrestres, arranjos de placas ou paleocorrentes. Também é necessária mais informação sobre possíveis tendências nos processos tafonômicos.
Várias tendências de diversidade podem ser interpretadas como resultado da expansão da atividade de inundação (ver Apêndice 1). Entre elas, estão aparências coordenadas (por exemplo, a explosão cambriana), aumentando a diversidade, a disparidade antes da diversidade e os desaparecimentos coordenados. A classificação estratigráfica precisa e consistente de fósseis em diferentes estratos é mais problemática. É necessário um mecanismo de classificação em larga escala para explicar a consistência da classificação em regiões geográficas de tamanho de continente. O Dilúvio pode fornecer esse mecanismo, mas os detalhes ainda precisam ser trabalhados. A tendência para aumentar a província também parece problemática para a teoria do dilúvio.
Os padrões morfológicos fornecem um saco misto para os criacionistas. A maioria dos padrões morfológicos é consistente com as expectativas criacionistas (ver Apêndice 2). Isso inclui estase morfológica em espécies fósseis, lacunas morfológicas entre espécies, lacunas sistemáticas entre táxons mais altos, estase com táxons mais altos, estase coordenada, falta de ancestrais e especialização morfológica onipresente. Muitas tendências de tamanho corporal podem ser esperadas em uma enchente, mas são necessários mais estudos para esclarecer os processos envolvidos. O aumento da complexidade pode ser um efeito secundário do aumento da terrestre entre os vertebrados. Os desafios mais significativos para o criacionismo a partir do registro fóssil são provavelmente a crescente semelhança com espécies modernas e a classificação estratigráfica de espécies em séries morfológicas. Essas tendências são as evidências fósseis mais importantes para a alternativa à teoria do dilúvio, a teoria da evolução. Um dos principais objetivos dos cientistas da criação deve ser o de fornecer explicações alternativas para séries morfológicas de fósseis. Algumas séries morfológicas foram ligadas a causas ecológicas e não evolutivas,[83] mas é necessário muito mais estudo nessa área.
CONCLUSÃO
O registro fóssil é um registro de destruição e morte. É o registro da história não direcionada, em que todas as espécies vivem por um tempo e depois se extinguem? Traça uma história evolutiva de ancestralidade comum, seleção natural e aprimoramento? Ou é um registro de destruição catastrófica mundial, projetada para servir como um lembrete dos efeitos do pecado? Somente a ciência não fornece uma resposta satisfatória, mas a Bíblia indica que a última interpretação é a correta. Os detalhes não são fornecidos e nenhuma teoria atual explica adequadamente todos os dados. Ninguém foi capaz de descobrir como juntar tudo. No entanto, comparando a Bíblia e o registro fóssil, podemos encontrar significado na coluna geológica. Atividade catastrófica e padrões globais, talvez as duas previsões mais importantes da teoria do Dilúvio, são claramente vistos no registro fóssil. O aparecimento repentino e abrupto de disparidade morfológica entre animais marinhos na "Explosão Cambriana" fala do início do dilúvio. O terrível poder destrutivo do dilúvio é visto em muitos grupos fósseis extintos. A falta de ancestrais nas rochas pré-cambrianas indica a criação separada de muitos grupos diferentes. A presença de lacunas morfológicas entre os táxons mais altos em todo o registro fóssil ilustra ainda mais esse ponto. A falta de ancestrais nas rochas pré-cambrianas indica a criação separada de muitos grupos diferentes. A presença de lacunas morfológicas entre os táxons mais altos em todo o registro fóssil ilustra ainda mais esse ponto. A falta de ancestrais nas rochas pré-cambrianas indica a criação separada de muitos grupos diferentes. A presença de lacunas morfológicas entre os táxons mais altos em todo o registro fóssil ilustra ainda mais esse ponto.
Nem todo mundo interpretará o registro dessa maneira. Mas aqueles que estão dispostos a testar suas idéias pela Bíblia podem ver o propósito divino no registro fóssil. Essa evidência afirma a realidade do propósito divino no presente e no futuro.
NOTAS FINAIS
[1] (a) Gould SJ. 1988. Tendências como mudanças de variação: uma nova tendência ao progresso e direcionalidade na evolução. Journal of Paleontology 62: 319-329. Veja também: (b) Gould SJ. 1996. casa cheia. NY: Harmony House; (c) Ruse M. 1993. Evolução e progresso. Tendências em Ecologia e Evolução 8: 55-59; (d) Raup DM, Gould SJ. 1974. Simulação estocástica e evolução da morfologia? Rumo a uma paleontologia nomotética. Zoology sistemático 23: 305-322.
[2] Por exemplo, consulte: (a) Benton MJ. 1993. Registro fóssil 2. Londres: Chapman e Hall; também, para vertebrados, consulte: (b) Carroll RL. 1988. Paleontologia e evolução de vertebrados. NY: Freeman; para invertebrados, ver: (c) Boardman RS, Cheetham AH, Rowell AJ, editores. 1987. Invertebrados fósseis. Blackwell. Boston.; para plantas, ver: (d) Stewart WN, Rothwell GW. 1993. Paleobotânica e a evolução das plantas. 2nd ed. Cambridge e NY: Cambridge University Press.
[3] Schopf JW. 1992. Procariontes proterozóicos: afinidades, distribuição geológica e tendências evolutivas. In: Schopf JW, Klein C, editores. A biosfera proterozóica. Cambridge: Cambridge University Press, p. 195-218.
[4]Um relato interessante da explosão cambriana está incluído em: (a) Gould SJ. 1989. Vida maravilhosa. NOVA IORQUE. WW Norton and Co. Outras referências incluem: (b) Conway Morris S. 1989. Faunas de Burgess Shale e a explosão cambriana. Science 246: 339-346; (c) Dzik J. 1993. Evolução inicial do metazoário e o significado de seu registro fóssil. Evolutionary Biology 27: 339-382; (d) Signor PW. 1990. A história geológica da diversidade. Revisão Anual de Ecology and Systematics 21: 509-539; (e) Valentine JW. 1994. Bilaterians pré-cambrianos atrasados: notas e clades. Anais da Academia Nacional de Ciências (EUA) 10: 190-194; f) Valentine JW. 1995. Por que nenhum novo filo depois do cambriano? Revisadas as hipóteses do genoma e do espaço. Palaios 10: 190-194; g) Valentine JW, Awramik SM, signor PW, Sadler PM. 1991. A explosão biológica na fronteira pré-cambriana-cambriana.
[5]Por exemplo, consulte: (a) Benton MJ. 1995. Diversificação e extinção na história da vida. Science 268: 5258; (b) Droser ML, Fortey RA, Xing Li. 1996. A radiação ordoviciana. American Scientist 84: 122-131; c) Doyle JA. 1977. Padrões de evolução nas angiospermas primitivas. In: Hallam A, editor. Padrões de evolução. Amsterdam e NY: Elsevier Scientific Publishing Co., p 501-546; (d) Erwin DH, Valentine JW, Sepkoski JJ. 1987. Um estudo comparativo de eventos de diversificação: o paleozóico inicial versus o mesozóico. Evolution 41: 1177-1186; (e) Feduccia A. 1995. Evolução explosiva em aves e mamíferos terciários. Science 267: 637-638; (f) Foote M. 1992. Registro paleozóico da diversidade morfológica nos equinodermes blastozoários. Anais da Academia Nacional de Ciências (EUA) 89: 7325-7329; (g) Foote M. 1994. Disparidade morfológica em crinóides Ordoviciano-Devonianos e saturação precoce do espaço morfológico. Paleobiology 20: 320-344; (h) Foote M. 1996. Sobre a probabilidade de ancestrais no registro fóssil. Paleobiology 22: 141-151; (i) Gould SJ, Gilinsky NL, RZ alemão. 1987. Assimetria de linhagens e a direção do tempo evolutivo. Science 236: 1437-1441; (j) Hunt AP, Lucas SG. 1991. Primeira aparição síncrona de dinossauros em todo o mundo durante o final do Triássico (Carniano tardio: Tuvaliano). Sociedade Geológica de Resumos Americanos com Programa 1991: A457; (k) Miller AI, Foote M. 1996. Calibrando a radiação ordoviciana da vida marinha: implicações para as tendências da diversidade fanerozóica. Paleobiology 22: 304-309; (l) Niklas KJ. 1994. Um passo gigante para a vida. História Natural 103 (6): 22-25; (m) Paul CRC. 1977. Evolução dos equinodermos primitivos. Em: Hallam, 123-158 (ver nota 5); n) Sepkoski JJ. 1981. Uma descrição analítica fatorial do registro fóssil marinho fanerozóico. Paleobiology 7: 36-53; (o) Zang, W-1. 1995. Estratigrafia de seqüência neoproterozóica inicial e bioestratigrafia de acritarca, Eastern Officer Basin, Austrália Meridional. Pesquisa pré-cambriana 74: 119-175.
[6]Extinções em massa são discutidas em dezenas de publicações. Alguns exemplos incluem: (a) Bardet N. 1994. Eventos de extinção entre répteis marinhos mesozóicos. Historical Biology 7: 313-324; (b) Benton 1995 (ver Nota 5); (c) Erwin DH. 1993. A grande crise paleozóica. NY: Columbia University Press; (d) Jablonski D. 1991. Extinções: uma perspectiva paleontológica. Science 253: 754-757; (e) JA longa. 1993. Eventos de extinção de vertebrados paleozóicos do início do meio. In: Long JA, editor. Bioestratigrafia e biogeografia de vertebrados paleozóicos. Baltimore: Johns Hopkins Press, p. 54-63; f) McGhee GR. 1996. A extinção em massa do Devoniano Tardio. NY: Columbia University Press; (g) Raup DM, Sepkoski JJ. 1982. Extinções em massa no registro fóssil marinho. Science 215: 1501-1503; h) Sepkoski JJ. 1993. Dez anos na biblioteca: novos dados confirmam padrões paleontológicos. Paleobiology 19: 43-51; i) Sepkoski JJ. 1994. Extinção e registro fóssil. Geotimes 39 (3): 15-17.
[7](a) Benton 1995 (ver nota 5); (b) Erwin, Valentine e Sepkoski 1987 (ver nota 5); (c) Knoll AH. 1984. Padrões de extinção no registro fóssil de plantas vasculares. In: Nitecki MH, editor. Extinção. Chicago e Londres: University of Chicago Press, p. 21-68; (d) Raup DM. 1976. Diversidade de espécies no Fanerozóico: uma tabulação. Paleobiology 2: 279-288 .; e) Rosenzweig ML. 1995. Diversidade de espécies no espaço e no tempo. Cambridge: Cambridge University Press; f) Sepkoski JJ, Sheehan PM. 1983. Diversificação, mudança de fauna e substituição da comunidade durante as radiações ordovicianas. In: Tevesz MJS, McCall PL, editores. Interações bióticas em comunidades bênticas recentes e fósseis. NY e Londres: Plenum Press, p 673-717; (g) Signor 1985. Tendências reais e aparentes da riqueza de espécies ao longo do tempo. In: Valentine JW, editor. Padrões de diversidade fanerozóica. Princeton, NJ: Princeton University Press, p. 129-150; h) Signor 1990 (ver nota 4); (i) Signor PW. 1994. Biodiversidade no tempo geológico. American Zoologist 34: 23-32.
[8] (a) Schopf JW. 1992. Padrões de diversidade de microfósseis proterozóicos: uma análise inicial e experimental. In: Schopf e Klein, p 529-552 (ver nota 3); (b) Walter MR, Grotzinger JP, Schopf JW. 1992. Stromatolites proterozóicos. In: Schopf e Klein, p 253-260 (ver Nota 3).
[9] Ver Gould 1989 (nota 4).
[10] Ver referências na nota 5.
[11] (a) Cox CB. 1974. Padrões paleodistributivos de vertebrados e deriva continental. Journal of Biogeography 1: 75-94 .; (b) Hutton N. 1992. Distribuição global de tetrápodes terrestres e aquáticos, e sua relevância para a posição das massas continentais. In: Chatterjee S, Hotton N, editores. Novos conceitos em tectônica global. Lubbock, TX: Texas Tech University Press, págs. 267-285; (c) Shubin NH, processa HD. 1991. Biogeografia dos primeiros tetrápodes continentais mesozóicos. Paleobiology 17: 214-230. Um padrão irregular de província está implícito em: (d) Wing SL, Sues HD. 1992. Ecossistemas terrestres mesozóicos e cenozóicos iniciais. In: Behrensmeyer AK, Damuth JD, DiMichele WA, Potts R, processa HD, Wing SL, editores. Ecossistemas terrestres através do tempo. Chicago e Londres: University of Chicago Press, p. 327-416.
[12] (a) McShea DW. 1996. Complexidade e evolução dos metazoários: existe uma tendência? Evolution 50: 477-492; b) Ruse 1993 (ver nota 1); (c) Valentine JW. 1994. Aumento da complexidade morfológica em metazoários. Paleobiology 20: 131-142.
[13] Schopf 1992, p. 195-218 (ver nota 3).
[14]a) Brett CE, Ivany LC, Schopf KM. 1996. Estase coordenada: uma visão geral. Paleogeografia, Paleoclimatologia, Paleoecologia 127: 1-20; (b) DiMichele WA, Hook RW. 1992. Ecossistemas terrestres paleozóicos. In: Behrensmeyer AK, Damuth JD, DiMichele WA, Potts R, processa HD, Wing SL, editores. Ecossistemas terrestres através do tempo. Chicago e Londres: University of Chicago Press, p. 205-325; (c) Eldredge N. 1996. Hierarquias na macroevolução. In: Jablonski D, Erwin DH, Lipps JH, editores. Paleobiologia evolutiva. Chicago e Londres: University of Chicago Press, p. 42-61; (d) Flynn LJ, Barry JC, Morgan ME, Pilbeam D, Jacobs LL, Lindsay EH. 1995. Linhagens de mamíferos Neogene Siwalik: longevidades de espécies, taxas de mudança e modos de especiação. Paleogeografia, Paleoclimatologia, Paleoecologia 115: 249-264; (e) Morris PJ, Ivany LC, Schopf KM, Brett CE. 1995. O desafio da estase paleoecológica: reavaliando fontes de estabilidade evolutiva. Anais da Academia Nacional de Ciências (EUA) 92: 11269-11273; (f) Tang CM, DJ Bottjer. 1996. Estase faunística a longo prazo sem coordenação evolutiva: paleocomunidades marinhas jurássicas bentônicas, Interior Ocidental, Estados Unidos. Geology 24: 815-818. Veja também as referências na Nota 15.
[15] (a) RJ fofo. 1984. Evidência paleontológica e evolução orgânica. In: Montagu A, editor. Ciência e criacionismo. Oxford e NY: Oxford University Press, p 255-281. Reproduzido do Journal of American Scientific Affiliation 24 (4, dezembro de 1972); (b) Gingerich PD. 1974. Registro estratigráfico de Early Eocene Hyopsodus e a geometria da filogenia de mamíferos. Nature 248: 107-109; c) Strahler AN. 1987. História da ciência e da terra: a controvérsia da evolução / criação. Buffalo, NY: Prometheus Books.
[16] (a) Bakker RT. 1985. Evolução por revolução. Science 85 (novembro): 72-80; (b) Eldredge N, Gould SJ. 1972. Equilíbrio pontuado: uma alternativa ao gradualismo filético. In: Schopf TJM, editor. Modelos de paleobiologia. San Francisco: Freeman, Cooper and Co., p. 82-115; (c) Gould SJ, Eldredge N. 1977. Equilíbrio pontuado: os tempos e o modo de evolução reconsiderados. Paleobiology 3: 115-151; (d) Sylvester-Bradley PC. 1977. Testes bioestratigráficos da teoria da evolução. In: Kauffman EG, Hazel JE, editores. Conceitos e métodos de bioestratigrafia. Stroudsburg, PA: Dowden, Hutchinson e Ross, p 41-63; (e) Heaton TH. 1993. Ischyromys, roedor do Oligoceno, das Grandes Planícies: substituição confundida com anagênese. Journal of Paleontology 67: 297-308.
[17] Schopf 1992, p. 195-218 (ver nota 3).
[18] (a) Sábio KP. 1991. Mudança de estase. Origins Research 13 (1): 20; b) Battson AL. 1994. Sobre a origem da estase por meio de processos naturais. Perspectivas sobre ciência e fé cristã 46: 230-241.
[19] (a) Brett, Ivany e Schopf 1996 (ver nota 14); (b) Ivany LC. 1997. Teste de estase coordenada na planície costeira do Golfo do Eoceno. Resumos da Sociedade Geológica com o Programa A404-405; (c) Morris, Ivany, Schopf e Brett 1995 (vide Nota 14). Muitos outros artigos sobre esse assunto estão em Brett et al. 1996.
[20] (a) Dommergues JL. 1990. Amonóides. In: McNamara KJ, editor. Tendências evolutivas. Tucson: University of Arizona Press, p. 162-187; (b) Eldredge e Gould 1972 (vide Nota 16); (c) Erwin, Valentine e Sepkoski 1987 (ver nota 5); (d) McNamara KJ. 1990. Epílogo. In: McNamara, págs. 347-350; (e) Williamson PG. 1981. Documentação paleontológica da especiação em moluscos cenozóicos da bacia do Turkana. Nature 293: 437-443; (f) ver também referências na Nota 11. Uma visão oposta é apresentada em: (g) Gingerich 1974 (vide Nota 15).
[21] Este ponto é discutido na página 191-194 de: (a) Denton M. 1986. Evolução: uma teoria em crise. Bethesda, MD: Adler e Adler; veja também (b) Carroll RL. 1992. A radiação primária dos vertebrados terrestres. Revisão Anual da Terra e das Ciências Planetárias 20: 45-84.
[22] Uma longa lista de reivindicações, muitas das quais foram reinterpretadas, é apresentada em: Cuffy 1984 (ver Nota 15). Para outras referências, consulte as Notas 24-27.
[23]a) Boucot AJ. 1976. As taxas de aumento de tamanho e de evolução filética. Nature 261: 694-695; (b) Cerdeno E, Nieto M. 1995. Alterações nas Rhinocerotidae da Europa Ocidental relacionadas a variações climáticas. Paleogeografia, Paleoclimatologia, Paleoecologia 114: 325-338; (c) Fortey RA, Owens RM. 1990. Trilobites. In: McNamara, p 121-142 (ver nota 20); (d) Jablonski D. 1997. Evolução do tamanho do corpo nos moluscos cretáceos e o status da regra de Cope. Nature 385: 250-252; (e) McKinney ML. 1986. Causação ecológica da heterocronia: um teste e implicações para a teoria da evolução. Paleobiology 12: 282-289; f) McKinney ML. 1990. Tendências na evolução do tamanho do corpo. In: McNamara, p. 75-118 (ver Nota 20); g) McNamara KJ. 1990. Equinóides. In: McNamara, p. 205-231 (ver Nota 20); (h) Mendelson CV, Schopf JW. 1992. Acritarcas proterozóicas e primitivas cambrianas. No: Schopf e Klein, p 219-232 (ver nota 3); (i) Morgan ME, Barry JC. 1991. Tendências do tamanho do corpo entre os herbívoros do Mioceno Médio e Tardio dos Siwaliks do norte do Paquistão. Journal of Vertebrate Paleontology 11 (Suppl): 48A, resumo 149.
[24] Benton MJ. 1990. Répteis. In: McNamara, p 279-300 (ver Nota 20).
[25]a) Clyde WC, Fisher DC. 1997. Comparando o ajuste de dados estratigráficos e morfológicos na análise filogenética. Paleobiology 23: 1-19; (b) Hitchin R, Benton MJ. 1997. Congruência entre parcimônia e estratigrafia: comparações de três índices. Paleobiology 23: 20-32; c) Huelsenbeck JP. 1994. Comparando o registro estratigráfico com estimativas de filogenia. Paleobiology 20: 470-483; (d) Norell MA, Novacek MJ. 1992. O registro fóssil e a evolução: comparando evidências cladísticas e paleontológicas para a história dos vertebrados. Science 255: 1690-1693; (e) Norell MA, Novacek MJ. 1992. Congruência entre padrões superposicionais e filogenéticos: comparação de padrões cladísticos com registros fósseis. Cladistics 8: 319-337; f) Padian K, Lindberg DR, Polly PD. 1994. Cladística e registro fóssil: os usos da história. Revisão Anual da Terra e Ciências Planetárias 22: 63-91; (g) Prado JL, Alberdi MT 1996. Uma análise cladística dos cavalos da tribo Equini. Paleontologia 39: 663-680; (h) Weishampel DB. 1996. Fósseis, filogenia e descoberta: um estudo cladístico da história de topologias de árvores e durações de linhagens fantasmas. Journal of Vertebrate Paleontology 16: 191-197. A falta de concordância estratigráfica é relatada em: (i) Antsey RL. 1990. Bryozoans. Em McNamara, p 232-252 (consulte a Nota 20).
[26] (a) Gould SJ. 1990. Especiação e classificação como fonte de tendências evolutivas ou "as coisas raramente são o que parecem". In: McNamara, p 3-27 (ver Nota 20); (b) MacFadden BJ. 1992. Cavalos fósseis. Cambridge e NY: Cambridge University Press.
[27] (a) Fraser NC, Sues HD, editores. 1994. À sombra dos dinossauros. Cambridge e NY: Cambridge University Press. Veja especialmente o Capítulo 6 (de Zhexi Luo) e o Capítulo 20 (de SG Lucas e AP Hunt); (b) Hotton N, MacLean PD, Roth JJ, Roth EC, editores. 1986. A ecologia e biologia de répteis semelhantes a mamíferos. Washington DC e Londres: Smithsonian Institution Press; (c) Kemp TS. 1982. Répteis semelhantes a mamíferos e a origem de mamíferos. Londres e NY: Academic Press.
[28] (a) Fastovsky DE, Weishampel DB. 1996. A evolução e extinção dos dinossauros. Cambridge e NY: Cambridge University Press; (b) Gatesy SM, disque KP. 1996. Módulos locomotores e a evolução do voo aviário. Evolution 50: 331-340. Para visão minoritária, veja: (c) Feduccia A. 1996. A origem e evolução das aves. New Haven e Londres: Yale University Press; (d) Martin LD. 1991. Aves mesozóicas e a origem das aves. In: Schultze HP, Trueb L, editores. Origens dos grupos superiores de tetrápodes. Ithaca e Londres: Comstock Publishing, p. 485-540.
[29] (a) Gingerich PD, Raza SM, Arif M, Anwar M, Zhou X. 1994. Nova baleia do Eoceno do Paquistão e a origem da natação de cetáceos. Nature 368: 844-847; (b) Thewissen JGM, Hussain ST, Arif M. 1994. Evidências fósseis da origem da locomoção aquática em baleias arqueocetas. Science 263: 210-212.
[30] (a) Landman NH. 1988. Heterocronia em amonites. In: McKinney ML, editor. Heterocronia na evolução: uma abordagem multidisciplinar. NY: Plenum, p 159-182; (b) Lidgard S, Jackson JBC. 1989. Crescimento em briozoários cheilostome incrustantes: I. Tendências evolutivas. Paleobiology 15: 255-282; (c) McNamara 1990 (vide nota 20); (d) Wagner PJ. 1996. Contrastando os padrões subjacentes de tendências ativas na evolução morfológica. Evolution 50: 990-1007.
[31] Este ponto pode ser confirmado através da análise dos dados em: a) Benton 1993 (ver nota 2). Veja também: (b) Horowitz AS, Pachut JF. 1994. Porcentagens de briozoários de Lyellian e registro fóssil da fauna recente de briozoários. Palaios 9: 500-505.
[32] Schopf 1992 (ver nota 3).
[33] Gould destaca esse ponto no livro Vida Maravilhosa (ver Nota 4).
[34] Cladistics não fornece um método para identificar ancestrais. Duas tentativas para fazê-lo são: (a) Alroy J. 1995. Análise de trilha contínua: um novo método filogenético e biogeográfico. Systematic Biology 44: 152-178; (b) Paul CRC. 1992. O reconhecimento dos antepassados. Historical Biology 6: 239-250. Um possível ancestral dos cães é identificado em: (c) Tedford RH, Taylor BE, Wang X. Exemplos mais problemáticos são: (d) Heinrich RE, Rose KD. 1995. Esqueleto parcial do carnívoro primitivo Miacis petilus do Eoceno Primitivo de Wyoming. Journal of Mammalogy 76: 148-162; (e) Rose KD. 1982. Esqueleto de Diacodexis , o mais antigo artiodáctilo conhecido. Science 216: 621-623; f) Rose KD. 1987. Adaptações de escalada no mamífero do início do Eoceno Chriacuse a origem do artiodactyla. Science 236: 314-316; g) Rose KD. 1996. Sobre a origem da ordem Artiodactyla. Anais da Academia Nacional de Ciências (EUA) 93: 1705-1709.
[35] (a) Benton MJ. 1996. Sobre a não prevalência de substituição competitiva na evolução dos tetrápodes. In: Jablonski, Erwin e Lipps, p 185-210 (ver Nota 14); (b) Droser ML, DJ Bottjer, Sheehan PM. 1997. Avaliando a arquitetura ecológica de grandes eventos na história fanerozóica da vida de invertebrados marinhos. Geology 25: 167-170; (c) Sepkoski e Sheehan 1983 (ver nota 7); (d) Sepkoski JJ, Miller AI. 1985. Faunas evolucionárias e distribuição de comunidades marinhas paleozóicas no espaço e no tempo. In: Valentine, p 153-190, (ver Nota 7); (e) Thayer CW. 1983. Perturbação biológica mediada por sedimentos e evolução dos bentos marinhos. In: Tevesz e McCall, p 479-625 (ver Nota 7); f) Vermeij GJ. 1987. Evolução e escalada. Princeton, NJ: Imprensa da Universidade de Princeton; (g) Wing and Sues 1992 (ver nota 11).
[36] a) Bambach RK. 1983. Utilização do espaço aeroespacial e guildas em comunidades marinhas através do Fanerozóico. In: Tevesz e McCall, p 719-746 (ver Nota 7); b) Bambach RK. 1985. Classes e variedade adaptativa: a ecologia da diversificação em faunas marinhas através do Fanerozóico. In: Valentine, p. 191-253 (ver Nota 7); c) Bambach RK, Bennington JB. 1996. As comunidades evoluem? Uma questão importante na paleoecologia evolutiva. In: Jablonski, Erwin e Lipps, p 123-160 (ver Nota 14); d) Sepkoski JJ. 1996. Competição em macroevolução: a dupla cunha revisitada. In: Jablonski, Erwin e Lipps, p 211-255 (ver Nota 14).
[37] (a) Conway Morris S, Whittington HB. 1979. Os animais do xisto de Burgess. Scientific American 241 (1): 122-133; b) Erwin 1993 (ver nota 6).
[38] (a) DiMichele WA, Aronson RB. 1992. A transição vegetacional Pennsylvanian-Permian: um análogo terrestre à hipótese onshore-offshore. Evolution 46: 807-824; (b) DiMichele e Hook 1992 (ver Nota 14); c) Niklas KJ, Tiffney BH, Knoll AH. 1985. Padrões na diversificação vascular de plantas terrestres: uma análise em nível de espécie. In: Valentine, p. 97-128 (consulte a Nota 7).
[39] Os únicos fósseis de vertebrados do Paleozóico inferior são as formas aquáticas. Eles são unidos por formas semi-aquáticas e de várzea no Paleozóico médio e superior, com um aumento das formas terrestres através do restante da coluna geológica. Por exemplo, veja: (a) DiMichele e Hook 1992 (veja Nota 14); (b) Olson, EC 1966. Evolução da comunidade e origem dos mamíferos. Ecology 47: 291-302. Para um ponto de vista diferente, consulte: (c) Padian K, Clemens WA. 1985. Diversidade de vertebrados terrestres: episódios e idéias. In: Valentine, p. 41-96 (ver Nota 7).
[40] Roth AA. 1992. Vida nas rochas profundas e registro fóssil profundo. Origens 19: 93-104.
[41] a) Erwin 1993 (ver nota 6); (b) Thayer 1983 (ver Nota 35); c) Vermeij 1987 (ver nota 35).
[42] (a) Cowen R. 1996. Locomoção e respiração em vertebrados respiradores do ar aquático. In: Jablonski, Erwin e Lipps, p. 337-353 (ver Nota 14); (b) Janis CM, Damuth J. 1990. Mammals. In: McNamara, p. 301-345 (ver nota 20); (c) Stucky RK. 1989. Evolução da diversidade de mamíferos terrestres na América do Norte durante o Cenozóico. Current Mammalogy 2: 375-432.
[43] Marsaglia KM, Klein GDeV. 1983. A paleogeografia dos sistemas deposicionais de tempestade paleozóica e mesozóica. Journal of Geology 91: 117-142.
[44] (a) Allison PA, Briggs DEG. 1993. Registro fóssil excepcional: distribuição da preservação de tecidos moles através do Fanerozóico. Geology 21: 527-530; b) Briggs DEG. 1991. Fósseis extraordinários. American Scientist 79: 130-141; (c) Butterfield NJ. 1996. Preservação do tipo xisto de Burgess de organismos cambrianos não mineralizantes e 'shelly' das montanhas MacKenzie, no noroeste do Canadá. Journal of Paleontology 70: 893-899.
[45] (a) DiMichele e Aronson 1992 (ver nota 38); Bottjer DJ, Jablonski D. 1988. Padrões paleoambientais na evolução de invertebrados marinhos bentônicos pós-paleozóicos. Palaios 3: 540-560; (b) Jablonski D, Sepkoski JJ, DJ Bottjer, Sheehan PM. 1983. Padrões onshore-offshore na evolução das comunidades de prateleiras fanerozóicas. Science 222: 1123-1125; (c) Jablonski D, DJ Bottjer. 1991. Padrões ambientais nas origens de táxons mais altos: o registro fóssil pós-paleozóico. Science 252: 1831-1833; (d) Miller AI. 1988. Transições espaço-temporais na bivalvia paleozóica: uma análise das assembléias fósseis norte-americanas. Biologia Histórica 1: 251-273; e) Sepkoski JJ. 1991. Um modelo de mudança onshore-offshore na diversidade da fauna. Paleobiology 17: 58-77.
[46] Behrensmeyer AK, Hook RW. 1992. Contextos paleoambientais e modos tafonômicos. In: Behrensmeyer, Damuth, DiMichele, Potts, Sues e Wing, p 15-136 (ver Nota 11).
[47] Ronov AB. 1982. A concha sedimentar da Terra (padrões quantitativos de sua estrutura, composições e evolução). International Geology Review 24: 1365-1388.
[48] (a) Behrensmeyer e Hook 1992 (ver nota 46); (b) Greenstein B. 1992. Viés tafonômico e a história evolutiva da família Cidaridae (Echinodermata: Echinoidea). Paleobiology 18: 50-79; (c) Kidder DL, Schubert JK, Erwin DH. 1997. Padrões seculares de silicificação esquelética de invertebrados. Resumos da Sociedade Geológica da América com o Programa A265; (d) Kidwell SM, Brenchley PJ. 1996. Evolução do registro fóssil: tendências de espessura nas acumulações esqueléticas marinhas e suas implicações. In: Jablonski, Erwin e Lipps, p 290-336 (ver Nota 14).
[49](a) Ausich WI, DJ Bottjer. 1985. Níveis fanerozóicos em comunidades de suspensão em substratos moles: implicações para a diversidade. In: Valentine, p. 255-274 (ver nota 7); (b) Durazzi JT, Stehli FG. 1972. Idade média genérica, gradiente de temperatura planetária e localização do pólo. Zoologia sistemática 21: 384-389; (c) Flessa KW, Jablonksi D. 1996. A geografia da rotatividade evolutiva: uma análise global dos bivalves existentes. In: Jablonski, Erwin e Lipps, p 376-397 (ver Nota 14); (d) Koch PL. 1986. Variação geográfica clinal em mamíferos: implicações para o estudo de cronoclinas. Paleobiology 12: 269-281; (e) JA longa. 1993. Eventos de extinção de vertebrados paleozóicos do início do meio. In: Long, p 54-63 (ver Nota 6); f) Raup DM, Stanley SM. 1978. Principles of paleontology. 2ª edição. São Francisco: Freeman, Capítulo 11, Evolução e registro fóssil; g) Rosenzweig ML. 1995 (ver nota 7); Signor 1990 (ver Nota 4); Signor 1994 (ver nota 7); Wing and Sues 1992 (ver nota 11).
[50] Raup e Gould 1974 (ver nota 1).
[51] (a) Cheetham AH. 1987. Tempo de evolução em um briozoário Neogene: as tendências em caracteres morfológicos únicos são enganosas? Paleobiology 13: 286-296; (b) Gould 1990 (vide Nota 26); c) Holland SM. 1995. A distribuição estratigráfica de fósseis. Paleobiology 21: 92-109; (d) McKee JK. 1995. Padrões de rotatividade e longevidade de espécies de grandes mamíferos do Plioceno e Pleistoceno tardios da África Austral: uma comparação de dados simulados e empíricos. Jornal de Biologia Teórica 172: 141-147.
[52](a) Aberhan M. 1993. Substituição da fauna no Jurássico Precoce do norte do Chile: implicações para a evolução nos ecossistemas mesozóicos da plataforma bentônica. Paleogeografia, Paleoclimatologia, Paleoecologia 103: 155-177; (b) Behrensmeyer e Hook 1992 (ver Nota 46); (c) Carlson JS. 1992. Tendências evolutivas no mecanismo articulado de braquiópodes. Paleobiology 18: 344-366; d) Cheema IU, Raza SM, Flynn LJ. 1989. Substituição radical de uma biota de pequenos mamíferos do Mioceno Precoce. Journal of Vertebrate Paleontology 9 (Suppl.): 16A; (e) DiMichele e Hook 1992 (ver Nota 14); f) McKinney ML. 1990. Classificação e análise de tendências evolutivas. In: McNamara, p. 28-58 (ver Nota 20); g) McNamara KJ. 1990. O papel da heterocronia nas tendências evolutivas. In: McNamara, p. 59-74 (ver Nota 20); (h) McNamara KJ. 1990. Equinóides. In: McNamara, p. 205-231 (ver Nota 20); (i) Miller AI. 1990. Bivalves. In: McNamara, p. 143-161 (ver Nota 20); (j) Simms MJ. 1990. Crinóides. In: McNamara, p 188-204 (consulte a Nota 20).
[53] Veja qualquer texto padrão sobre geologia histórica ou paleontologia, por exemplo: (a) Poort JM, Carlson RJ. 1997. Geologia histórica: interpretações e aplicações. 5ª edição. Englewood Cliffs, NJ: Prentice-Hall; (b) Rogers JJW 1993. Uma história da Terra. Cambridge: Cambridge University Press.
[54] (a) Van Valen L. 1973. Uma nova lei evolucionária. Teoria Evolucionária 1: 1-30; b) Vermeij 1987 (ver nota 35).
[55] Bengtson S, Lipps JH. 1992. Introdução. In: Schopf e Klein, p. 427-428 (ver Nota 3).
[56] Quando pode ser verificado, o registro fóssil não parece tão incompleto. Veja: (a) Foote 1996 (veja Nota 5); (b) Foote M, Raup DM. 1996. Preservação fóssil e faixas estratigráficas de táxons. Paleobiology 22: 121-140; (c) Valentine JW. 1989. Quão bom foi o registro fóssil? Pistas do Pleistoceno da Califórnia. Paleobiology 15: 83-94.
[57] (a) Gould 1988 (ver Nota 1); (b) Gould 1990 (vide Nota 26).
[58](a) Marca L. 1997. Fé, razão e história da terra: um paradigma da terra e das origens biológicas do design inteligente. Berrien Springs, MI: Andrews University Press; (b) Clark HW. 1946. O novo diluvialismo. Angwin, CA: Publicações de Ciências; (c) Clark HW. 1968. Fósseis, inundações e incêndios. Escondido, CA: Imagens ao ar livre; (d) Coffin HG. 1969. Criação: acidente ou design? Washington DC: Review and Herald Publishing Assn .; (e) caixão HG. 1983. Origem por design. Hagerstown, MD: Review and Herald Publishing Assn .; f) Oard MJ. 1997. A extinção dos dinossauros. Diário Técnico da Criação Ex Nihilo 11: 137-154; (g) Snelling AA. 1991. Geologia criacionista: onde se encaixam os estratos pré-cambrianos? Diário Técnico da Criação Ex Nihilo 5 (2): 154-175; (h) Whitcomb JC, Morris HM. 1961. O dilúvio de Gênesis. Phillipsburg, NJ: Publicação Presbiteriana e Reformada; (i) Wise K. 1989. Estilo de criação Punc Eq. Origens 16: 11-24; (j) Wise 1991 (ver nota 18); (k) KP sábio. 1992. Algumas reflexões sobre o registro fóssil pré-cambriano. Diário Técnico da Criação Ex Nihilo 6 (1): 67-71; (l) Sábio KP. 1994.Australopithecus ramidus e o registro fóssil. Diário Técnico da Criação Ex Nihilo 8: 160-165; (m) Sábio KP. 1995. Em direção a um entendimento criacionista de 'formas transitórias'. Diário Técnico da Criação Ex Nihilo 9 (2): 216-222; (n) KP sábio. 1996. Convenção Norte-Americana de Paleontologia 96. Creation Ex Nihilo Technical Journal 10: 315-321; (o) Woodmorappe J. 1983. Um tratado diluviológico sobre a separação estratigráfica de fósseis. Creation Research Society Quarterly 20: 133-185.
[59] (a) Clark 1946 (ver nota 58); (b) caixão de 1983 (ver nota 58); (c) Sábio de 1996 (ver nota 58).
[60] Sábio de 1996 (ver nota 58).
[61] Marca 1997, p 242-243, 299 (ver nota 58).
[62] (a) Battson 1994 (ver nota 18); b) Marsh FL. 1941. Biologia fundamental. Lincoln, NE: publicado pelo autor; (c) Wise 1989 (ver Nota 58); (d) Sábio 1991 (ver Nota 18).
[63] Sábio de 1989 (ver nota 58).
[64] Whitcomb e Morris 1961 (ver nota 58).
[65] Sábio de 1989 (ver nota 58).
[66] Ibidem.
[67] (a) Clark 1946 (ver nota 58); (b) Clark 1968 (ver Nota 58); (c) Whitcomb e Morris 1961 (ver Nota 58).
[68] Whitcomb e Morris 1961 (ver nota 58).
[69] (a) Clark 1946 (ver nota 58); (b) Clark 1968 (ver nota 58).
[70] Acredita-se que fósseis de bactérias fotossintéticas estejam presentes nos estratos pré-cambrianos, conforme observado em: (a) Schopf 1992 (consulte a Nota 3). No entanto, sua presença nos estromatólitos proterozóicos "ainda não foi adequadamente documentada", de acordo com: (b) Walter, Grotzinger e Schopf, 1992 (vide Nota 8).
[71] Snelling 1991 (ver nota 58).
[72] Wise 1992 (ver nota 58).
[73] Roth 1992 (ver nota 40).
[74] (a) Davies DJ, Powell EN, Stanton RJ. 1989. Taxas relativas de dissolução de conchas e acúmulo líquido de sedimentos um comentário: os leitos de conchas podem se formar pelo acúmulo gradual de detritos biogênicos no fundo do mar? Lethaia 22: 207-212; (b) Kidwell SM, Brenchley PJ. 1994. Padrões na acumulação bioclástica através do Fanerozóico: mudanças na entrada ou na destruição? Geology 22: 1139-1143; (c) Kidwell SM, Flessa KW. 1996. A qualidade do registro fóssil: Populações, espécies e comunidades. Revisão Anual da Terra e das Ciências Planetárias 24: 433-464. Veja também a Nota 41.
[75] Woodmorappe 1983 (ver nota 58).
[76] Marca 1997 (ver nota 58).
[77] Woodmorappe 1983 (ver nota 58).
[78] (a) Olson 1966 (ver nota 39); (b) Sábio de 1995 (ver Nota 58). Observe que Olson não é um criacionista.
[79] Mehlert AW. 1993. A origem dos mamíferos: um estudo de alguns fósseis importantes. Creation Ex Nihilo Technical Journal 7: 122-139.
[80] Sábio de 1995 (ver nota 58).
[81] (a) Marca 1997 (ver nota 58); (b) Sábio de 1989 (ver Nota 58).
[82] (a) Holt RD. 1996. Evidência de um limite de inundação cainozóica tardia / pós-inundação. Diário Técnico da Criação Ex Nihilo 19: 128-167; (b) Oard 1996 (vide Nota 58); (c) Oard MJ. 1997. A extinção dos dinossauros. Diário Técnico da Criação Ex Nihilo 11: 137-154.
[83] Explicações ecológicas de tendências morfológicas não são inconsistentes com a teoria da evolução, mas fornecem uma possível explicação que é consistente com o Dilúvio. Veja as referências na Nota 41 para exemplos.







